Mechanisms of Meat Curing – From Hoagland in 1914 to Pegg and Shahidi in 2000.
By: Eben van Tonder
14 August 2016

BACKGROUND AND OVERVIEW
Managing a meat curing operation in Cape Town at Woodys Consumer Brands (Pty) Ltd., the need exist to have a thorough understanding of meat curing mechanisms to ensure that conditions exist to optimise cured colour development, limit bacterial growth and deliver good product flavour and taste.
In the next three articles we look at colour development. This first article sets the historical context by reviewing the 1914 landmark article by Hoagland; we briefly outline the current understanding of cured colour development from the work of Pegg and Shahidi and we overview one mechanism that has recently been described. Overall, we focus on the importance of nitric oxide (NO) in cured colour development for both fresh and cooked cured meat.
INTRODUCTION
The formation of cured meat colour takes place “by the reaction of nitrite with the natural meat pigment myoglobin to form dinitrosyl ferrochrome (DNFH). The pigment, which gives meat its characteristic cured-meat colour, is formed from the meat pigment myoglobin, which consists of an iron porphyrin complex, the heme group, attached to the protein globin. In the presence of nitrite, the bright red nitrosomyoglobin is formed, in which the H2O in the axial position on the heme iron is replaced by nitric oxide (NO). The NO is formed from nitrite by the natural reducing activity of the muscle tissue, which is accelerated by the addition of reductants such as ascorbic acid. In heatprocessed cured meat, the globin has been split off to a heat-stable pink pigment, nitrosyl hemochromogen.” (Soltanizadeh, N., Kadivar, M.. 2012)
This understanding of curing developed over many years with input from a variety of scientists. (The Fathers of Modern Meat Curing) One of these influential minds was Ralph Hoagland. His brilliance is seen in his academic work that helped to shape the meat curing industry. He had wide appeal in academic and industry circles as well as the popular press. He contributed immensely to the developing sciences of nutrition and meat processing with a special interest in pork processing and pork nutrition.
He was the Senior Biochemist, Biochemie Division, Bureau of Animal Industry, United States Department of Agriculture in Chicago who was, at this time, one of the curing centers of the world along with Denmark and Calne, in the United Kingdom where the Harris operation started.
He served as the department head of the Minnesota College of Agriculture (part of the University of Minnesota), appointed in 1909. The College of Agriculture later became the College of Biological Sciences. (http://cbs.umn.edu/ and The Bismarck Tribune; 1912)
In 1908 he published results obtained upon studying the action of saltpeter upon the colour of meat and “found that the value of this agent in the curing of meats depends upon its reduction to nitrites and nitric oxid, with the consequent production of NO-hemoglobin, to which compound the red color of salted meats is due.” He found that “saltpeter, as such, [had] no value as a flesh-color preservative.” (Hoagland, R. 1914)
In 1914 he published, Colouring Matter of Raw and Cooked Salted Meat. Reviewing this article has two important objectives.
1. It shows what was understood by 1914 about meat curing and colour formation in particular. This has important implication for determining an accurate chronology of developments around the direct addition of nitrite to curing brines, such as the invention of Praganda in Prague in 1915 and later, the introduction of Prague Salt in Chicago (The Naming of Prague Salt) where Hoagland worked for a time.
2. It is a novel way for an introduction into meat curing mechanisms and shows the progression in our understanding.
I interject the thoughts of Hoagland from 1914 with quotes on our current understanding by two of the leading scientists on the subject namely Ronald B. Pegg and Fereidoon Shahidi with quotes from their 2000 publication, Nitrite Curing of Meat.
Ronald Pegg is currently a professor at the Department of Food Science & Technology, University of Georgia. A great piece appeared about him in FST News (from the University of Georgia Department of Food Science and Technology). “He is a researcher who feels equally at home in the classroom and the laboratory. In addition to inspiring students with the chemistry of chocolate and coffee, he’s become one of the nation’s most sought-after experts on the nutrient content of food and the bioactive compounds that make blueberries, peanuts and other nutritionally dense superfoods so “super.” Pegg joined the faculty of UGA in 2006. He immediately saw the need for a more hands-on, practical approach to teaching food chemistry. His work with students has earned him Food Science and Technology Outstanding Undergraduate and Graduate Professor awards five times. Pegg has received a major teaching honor from his department, the college or the university every year since 2007.” “In addition to his time in the classroom, Pegg has received accolades from producer groups for his research into bioactive chemistry and the health benefits of pecans, peanuts, peaches and other crops.” (http://www.foodscience.caes.uga.edu/)
His research and publishing partner in Nitrite Curing of Meat is Fereidoon Shahidi. He is a university research professor at the Department of Biochemistry, Memorial University of Newfoundland St. John’s, Canada. This monumental food scientist “has received numerous awards, including the 2005 Stephen Chang Award from the Institute of Food Technologists, for his outstanding contributions to science and technology. Between 1996 and 2006, Shahidi was the most published and most frequently cited scientist in the area of food, nutrition, and agricultural science as listed by the ISI.” (wikipedia.org/wiki/Fereidoon_Shahidi)
THE COLOUR OF FRESH MEAT
Hoagland starts with the colour pigment of fresh meat, oxyhemoglobin. The word itself tells us what it is. Oxy is oxygen, connected to hem which is hamatin or the colouring group and globin, the protein. In Oxyhemoglobin, ogygen is connected to “hemoglobin, which is the protein molecule in red blood cells that carries oxygen from the lungs to the body’s tissues and returns carbon dioxide from the tissues back to the lungs.” (medicinenet)
Hoagland states that oxyhemoglobin, is “part of which is one of the constituents of the blood remaining in the tissues, while the remainder is a normal constituent of the muscles,” and “responsible for the red color of fresh lean meat, such as beef, pork, and mutton.” (Hoagland, R. 1914) Today we know that the colour of fresh lean meat is due to myoglobin, “the pigment in muscle that carries oxygen” (medicinenet), as opposed to protein in the blood.
The reason for using hemoglobin, instead, may have been “a matter of convenience” and “a matter of necessity since myoglobin was not isolated and purified until 1932,” (Theorell, 1932) a full 18 years after Hoagland published. “In spite of the differences between hemoglobin and myoglobin, Urbain and Jensen (1940) considered the properties of hemoglobin and its derivatives sufficiently like those of myoglobin to allow the use of hemoglobin in studies of meat pigments.” (Cole, Morton Sylvan, 1961: 2)
Despite the fact that it is oxymyoglobin that is responsible for the bright red colour of fresh meat, we follow his arguments using oxyhemoglobin since the same mechanisms of colour development applies in both proteins. Pegg and Shahidi uses myoglobin.
Our current understanding: Oxymyoglobin (Mb , bright red,
, bright red,  – ferrous state)
– ferrous state)
Oxymyoglobin is the result of myoglobin’s affinity for and it results in a bright red bloom within minutes of fresh meat’s exposed to air. The reaction is rapid and reversible. The continued red bloom depends on a “continuing supply of .” (Pegg, R. B and Shahidi, F; 2000: 31) This is “because the enzymes involved in oxidative metabolism rapidly use the available .” (Pegg, R. B and Shahidi, F; 2000: 31)
“With time, the small layer of oxymyoglobin present on the surface of the meat propagates downward, but the depth to which diffuses depends on several factors, such as the activity of oxygen-utilizing enzymes (i.e., consumption rate of the meat), temperature, pH, and external pressure. In other words, as air diffuses inward, an and a color gradient are established throughout the meat. Muscles differ in their rates of enzyme activity which, in turn, regulate the amount of available in the outermost layers of tissue. As the pH and temperature of the tissue increase, enzymes become more active and the content is reduced. Consequently, maintaining the temperature of the meat near freezing point minimizes the rate of enzyme activity and the utilization and helps maintain a bright red color for the maximum possible time.” (Pegg, R. B and Shahidi, F; 2000: 31)
THE COLOUR OF CURED MEAT
Generally, Hoagland saw the cured colour of meat as “the same color as the fresh meat.” (Hoagland, R. 1914) There is a difference between the cured colour of fresh meat and the cured colour of cured-cooked meat. He recognised this difference and said that “the red color is not destroyed on cooking, but rather it is intensified.” (Hoagland, R. 1914)
The nature of these two different kinds of colour is the subject of his article, “undertaken for the purpose of obtaining more complete information concerning the color of raw and cooked salted meats.” (Hoagland, R. 1914)
HISTORICAL BACKGROUND
In his historical summary, he lists the following developments that lead up to his own work.
-> Weiler and Riegel
“Weiler and Riegel (1897), in the examination of a number of samples of American sausages, obtained a red coloring matter on extracting the samples with alcohol and other solvents, which color they concluded to be in some manner due to the action of the salts used in curing upon the natural color of the meat. On account of similarity of spectra, this color was considered to be methemoglobin.” (Hoagland, R. 1914)
Our current understanding: metmyoglobin (metMb, brown,  ferric state)
ferric state)
Methemoglobin and metmyoglobin actually is the brown colour of meat which develops after meat has been standing for some time. Myoglobin exists within the interior of meat and has a purple-red colour. “This is the colour of Myoglobin” (Pegg, R. B and Shahidi, F; 2000: 31) Reductants generated within a cell by enzyme activity prevents the meat from turning brown, until this is no longer available. The heme iron (in the ferrous state – ) is oxidized to the ferric state () . (Pegg, R. B and Shahidi, F; 2000: 31)
It is generated as follows. The superoxide anion ( ) is removed from the hematin. A water molecule is added. This gives a high-spin ferric hematin. “The ferric ion, unlike its ferrous counterpart, has a high nuclear charge and does not engage in strong
) is removed from the hematin. A water molecule is added. This gives a high-spin ferric hematin. “The ferric ion, unlike its ferrous counterpart, has a high nuclear charge and does not engage in strong 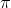 bonding. Therefore, metmyoglobin is unable to form an oxygen adduct. (Pegg, R. B and Shahidi, F; 2000: 31)
bonding. Therefore, metmyoglobin is unable to form an oxygen adduct. (Pegg, R. B and Shahidi, F; 2000: 31)
-> Lehmann and Kisskalt
Lehmann (1899) identified nitrite as responsible for the red colour of meat and not nitrate. Kisskalt (1899) confirmed this and noted that “if the meat was first allowed to stand several days in contact with saltpeter and then boiled, the red color appeared” (Hoagland, R. 1914)
-> John Scott Haldane
John Scott Haldane (1901) made several important observations after an extensive study of the colour of cooked salted meat.
He attributed the colour of cooked salted meat “to the presence of the nitric oxide hemochromogen” (reduced hematin; Fe in reduced ferrous state, ; obtained by boiling oxymyoglobin/ oxyhemoglobin with a reducing agent). (Hoagland, R. 1914) He correctly concluded that nitric oxide hemochromogen is “resulting from the reduction of the coloring matter of the uncooked meat, nitric-oxid hemoglobin (NO-hemoglobin).” Hemochrome can be any of a number of complexes with the iron-porphyrin complex with one or two basic ligands (normally amines).
The terms nitric oxide hemochromogen, nytrosomyochrome, nitrosyl hemochrome, nitric oxide hemochrome, nitric oxide denatured globin hemochromogen, denatured globin nitric oxide ferrohemochrome, pigment of cured, heated meat, all as synonyms to refer to the same thing. (ICMSF; 1980: 140) Chromogen is a substance which can be easily converted into dye or other coloured compound for example through oxidation. Since the 1940’s, the term “hemochrome” (hem and chrome) has been used instead of “hemochromogen” and “parahematin.” “The term “hemochromogen” is associated historically with an erroneous conception of one of these substances as the colored component of hemoglobin. These compounds are in any case not “chromogens” in the chemical sense, i.e., ieuco compounds. The new term has the additional advantage of greater brevity.” (Lemberg, R. and Legge, J. W.; 1949: 165)
Linossier was the first to describe it and produced it by passing nitric oxide through hematin. (Haldane, J. S.. 1901) After careful study and observation, Haldene drew the following brilliant conclusions.
1. “The red colour of cooked salt meat is due to the presence of NO-haemochromogen.” (Haldane, J. S.. 1901)
2. “The NO-haemochromogen is produced by the decomposition by heat of NO-haemoglobin, to which the red colour of unsalted meat is due.” (Haldane, J. S.. 1901)
3. “The NO-haemoglobin is formed by the action of nitrite on haemoglobin in the absence of oxygen, and in presence of reducing agents.” (Haldane, J. S.. 1901)
4. “The nitrite is formed by reduction within the raw meat of the nitre used in salting.” (Haldane, J. S.. 1901)
5. “The nitrite is destroyed by prolonged cooking.” (Haldane, J. S.. 1901)
Our current understanding: nitric oxide hemochrome (Cooked Cured Meats – one nitric oxide molecule per heme).
When heated, NO-myoglobin (nitrosyl myoglobin) is transformed to nitrosyl myocromogen, which is denatured NO-myochromogen. This happens upon thermal processing. The globin unfolds (denatures); the iron atom comes loose from the globin; the unfolded globin folds itself around the heme functional part (moiety) which is the iron-porphyrin complex. This brings about the characteristic reddish-pinkish colour of cooked cured meat. (Pegg, R. B and Shahidi, F; 2000: 42)
By way of application, note that “there is a direct relationship between the concentration of NO-myoglobin in the muscle and the intensity of the cured colour” and NOT the nitrite level. “When muscle tissue are cured with equivalent amounts of nitrite, a more intense cured meat colour is produced in,” for example, corned beef as opposed to ham. “The addition of excess nitrite to that required to fix the pigment does not increase the intensity of the cured meat colour.” (Pegg, R. B and Shahidi, F; 2000: 42) This being the case, it is also true that if the concentration of nitrite and therefore nitric oxide formation is to low, that it will impact colour development.
He mentions Orlow (1903) who stated that “the red color of sausages is due to the action upon the color of the fresh meat of the nitrites resulting from the reduction of the saltpeter used in the process of manufacture.” (Hoagland, R. 1914)
“Humphrey Davy in 1812 (cited by Hermann, 1865) and Hoppe-Seyler (1864) noted the action of nitric oxid upon hemoglobin, but it appears that Hermann (1865) was the first to furnish us with much information as to the properties of this derivative of hemoglobin. He prepared NO-hemoglobin by first passing hydrogen through dog’s blood until spectroscopic examination showed that all of the oxyhemoglobin had been reduced to hemoglobin, then saturating the blood with pure nitric oxid prepared from copper and nitric acid, and finally again passing hydrogen through the blood to remove all traces of free nitric oxid.” (Hoagland, R. 1914)
By the time of publishing this article in 1914, he notes that NO-hemoglobin was mentioned very briefly in most of the texts on physiological or organic chemistry as being a hemoglobin derivative of “but little practical importance.” “Abderhalden (1911) and Cohnheim (1911), however, describe this compound quite fully.” (Hoagland, R. 1914)
Hoagland conducted several further experiments with NO-hemoglobin and outlined it in his 1914 paper.
COLOUR OF FRESH, CURED MEAT
He first deals with the Colour of Uncooked Salted Meats. “To a sample of finely ground fresh beef was added 0.2 per cent of potassium nitrate, and the material was placed in a refrigerated room at a temperature of 34 deg F (1 deg C) for seven days. At the end of that period the meat had a bright-red color, but gave evidences of incipient putrefaction.” (Hoagland, R. 1914) He did the same by curing the meat with nitrite. He correctly concluded that the colour of fresh meat, cured with nitrite, is due to NO-hemoglobin. (Hoagland, R. 1914)
Our current understanding: nitric oxide myoglobin (NOMb, red, ).
“When nitrite is added to comminuted meat, the meat turns brown because nitrite acts as a strong heme oxidant. The oxidizing capacity of nitrite increases as the pH of meat decreases, but nitrite itself may also partly be oxidized to nitrate during curing and storage. Myoglobin and  are oxidized to metMb by nitrite. The ion itself can be reduced to
are oxidized to metMb by nitrite. The ion itself can be reduced to  . These products can combine with one another to form an intermediate pigment, nitrosylmetmyogloboin (
. These products can combine with one another to form an intermediate pigment, nitrosylmetmyogloboin ( ).” (Pegg, R. B and Shahidi, F; 2000: 40)
).” (Pegg, R. B and Shahidi, F; 2000: 40)

“Nitrosylmetmyoglobin is unstable. It auto-reduces with time and in the presence of endogenous and exogenous reductants in the postmortem muscle tissue to the corresponding relatively stable Fe(II) form, nitrosylmyoglobin (NOMb).” (Pegg, R. B and Shahidi, F; 2000: 40)
A new suggestion was proposed as a mechanism for the meat curing process by Killday et al. (1988)

“They suggested that is more adequately described as an imidazole-centered protein radical. This radical undergoes autoreduction yielding NOMb, and lacking exogenous reductants, reducing groups within the protein can donate electrons to the imidazole radical.” (Pegg, R. B and Shahidi, F; 2000: 40)
An interesting study by Corforth et al. (1998) strengthened the mechanism posed by Killday et al. (1988). “Cornforth and co-workers examined the relative contribution of CO and  towards pink ring formation in gas oven cooked beef roast and turkey rolls. Data showed that pinking was not evident with up to 149 ppm of CO or 5 ppm of NO present in the burning gases; however, as little as 0.4 and 2.5ppm of
towards pink ring formation in gas oven cooked beef roast and turkey rolls. Data showed that pinking was not evident with up to 149 ppm of CO or 5 ppm of NO present in the burning gases; however, as little as 0.4 and 2.5ppm of  was sufficient to cause pinking of the turkey and beef products, respectively. Cornforth et al. (1998) proposed that pinking previously attributed to CO and NO gas in ovens is instead due to which has much greater reactivity than NO with moisture at the surface of meats. Their argument was predicated on the fact that NO has a low water solubility unlike that of . Therefore on the basis of this consideration, NO would be an unlikely candidate to cause pink ring, since at the low levels typical of gas ovens or smokehouses, NO would be unable to enter the aqueous meat system in sufficient quantity to cause pink ring at depths up to 1 cm from the surface. On the other hand, reacts readily with water to produce nitrous and nitric acid.” (Pegg, R. B and Shahidi, F; 2000: 40, 42)
was sufficient to cause pinking of the turkey and beef products, respectively. Cornforth et al. (1998) proposed that pinking previously attributed to CO and NO gas in ovens is instead due to which has much greater reactivity than NO with moisture at the surface of meats. Their argument was predicated on the fact that NO has a low water solubility unlike that of . Therefore on the basis of this consideration, NO would be an unlikely candidate to cause pink ring, since at the low levels typical of gas ovens or smokehouses, NO would be unable to enter the aqueous meat system in sufficient quantity to cause pink ring at depths up to 1 cm from the surface. On the other hand, reacts readily with water to produce nitrous and nitric acid.” (Pegg, R. B and Shahidi, F; 2000: 40, 42)

“Nitrous acid produced at meat surfaces would be free to diffuse inwards, where endogenous or exogenous meat reductants, including Mb itself may regenerate NO. Nitric oxide binds to MetMb followed by rapid autoreduction to NOMb as suggested by Killday et al. (1988).” (Pegg, R. B and Shahidi, F; 2000: 42)



NOMb is therefore responsible for the characteristic red colour of fresh cured meat before thermal processing. The NOMb pigment can be produced by the direct action of NO on a deoxygenated solution of Mb, but in conventional curing, it arises from the action of nitrite, as stated above. (Pegg, R. B and Shahidi, F; 2000: 42)
Hoagland’s conclusion in his 1914 article is, however, limited to NO formation and its role in cured colour formation. He states that “the evidence is ample to show that the action of saltpeter in the curing of meats is primarily to cause the formation of NO-hemoglobin ; but it is very possible that under certain conditions of manufacture or processing to which salted meats are subject, the NO-hemoglobin may undergo changes.”
COLOUR OF COOKED, CURED MEAT
“Haldane has shown that the red color of cooked salted meats is due to the presence of NO-hemochromogen, a reduction product of NO-hemoglobin to which the color of uncooked salted meats is due.”… “While Haldane’s work seems to show clearly that the color of cooked salted meats is due to NO-hemochromogen, it has seemed desirable to study the subject further and to determine especially if the NO-hemoglobin of uncooked meats be reduced to NO-hemochromogen under other conditions than by cooking. The fact that in the examination of certain uncooked salted meats a coloring matter had been obtained similar to NO-hemoglobin yet not possessing all of the properties of that compound, as has already been noted, led the writer to believe that the coloring matter of some uncooked salted meats might be due, in part at least, to NO-hemochromogen. NO-hemochromogen is but briefly mentioned in the literature. The compound is described by Linossier (1887), Haldane (1901), and by Abderhalden (1911).” (Hoagland, R. 1914)
“The structural relation between NO-hemoglobin and NO-hemochromogen is simple. NO-hemoglobin is a molecular combination of nitric oxid and hemoglobin—the latter compound consisting of the proteid group, globin, on one hand, and the coloring group, hemochromogen, on the other. NO-hemoglobin and NO-hemochromogen differ from each other simply in that one contains the proteid group, globin, while the other does not. Apparently, then, a method of treatment which would split off the globin group from NO-hemoglobin should result in the production of NO-hemochromogen, provided, of course, that the procedure did not in turn change or destroy the NO-hemochromogen produced. As has already been noted by Haldane, it was found that when a solution of NO-hemoglobin was heated to boiling, a brick-red precipitate formed, in contrast to the dark-brown precipitate which formed on heating a solution of oxyhemoglobin or of blood. The brick-red precipitate was filtered off and was then extracted with alcohol, which gave a lightred colored extract showing a spectrum with a fairly heavy band just at the right of the D line. This spectrum corresponds with that of NO-hemochromogen. On standing, the color of the extract faded rapidly.” (Hoagland, R. 1914)
“The evidence seems to show very clearly that the color of cooked salted meats is due to the NO-hemochromogen resulting from the reduction of the NO-hemoglobin of the raw salted meats on boiling.” (Hoagland, R. 1914)
“It is very probable that in the case of meats which have been cured with saltpeter or of meat food products in which saltpeter has been used in the process of manufacture, the reduction of NO-hemoglobin to NO-hemochromogen takes place to a greater or lesser degree, depending upon conditions of manufacture and storage. The two compounds are so closely allied that their differentiation in one and the same product is not a matter of great importance.” (Hoagland, R. 1914)
Our current understanding: Nitrosylmyochromogen or nitrosylprotoheme.
Upon thermal processing, globin denatures and detaches itself from the iron atom and surrounds the hem moiety. Nitrosylmyochromogen or nitrosylprotoheme is the pigment formed upon cooking , and it confers the characteristic pink colour to cooked cured meats.” (Pegg, R. B and Shahidi, F; 2000: 44)
“Although the Cooked Cured Meat Pigment (CCMP) is a heat-stable NO hemochrome as evident by the fact that it doesnt undergo further colour change upon additional thermal processing, it is susceptible to photodissociation. Furthermolre in the presence of oxygen, CCMP’s stability is limited by the rate of loss of NO.

This effect is important if cured meats are displayed under strong fluorescent lighting while they are also exposed to air. Under these conditions, the surface colour of cured meat will fade in a few hours, whereas under identical conditions, fresh meat will hold its colour for a few days.” “A brownish-gray colour develops on the exposed meat surface during colour fading; this pigment, sometimes called hemichrome, has its heme group in the ferric state. The most effective way of preventing light fading is to exclude contact with the cured meat surfaces. It is routinely accomplished by vacuum packaging the meat in impermiable films. If is absent from the package, NO cleaved from the heme moieties by light cannot be oxidized and can recombine with the heme.” (Pegg, R. B and Shahidi, F; 2000: 44)
CONCLUSION
Hoagland and other researchers from that period laid the foundation to much of our current understanding of meat curing by drawing a distinction between fresh cured meat colour and cooked cured colour. The first detailed mechanism in the development of cured meat colour that started to emerge was through the action of nitric oxide. Pegg and Shahidi stated in 2000 that “to form cured meat pigment, two reduction steps are necessary. The first reduction of nitrite to NO and the second is conversion of NOmetMB to NOMb.” (Pegg, B. R. and Shahidi, F.; 2000: 44, 45)
An interesting side note. Hoagland wondered if it is possible to produce the cooked cured colour of meat in another way than curing with nitrite and heat treatment. Pegg and Shahidi has dedicated much work along similar lines – to identify a curing system that will replace nitrite curing. In meat curing, this has always been the holy grail which on the one hand will in all likelihood remain an unattainable concept and on the other hand, as our understanding of nitrite grows, will be deemed unnecessary.
The chemical reaction sequence from nitrite to NO, leading to the formation of NOMb will be described in the next article.
References:
The Bismarck Tribune (Bismarck, North Dakota); 10 July 1912; page 2.
Cole, Morton Sylvan, “Relation of sulfhydryl groups to the fading of cured meat ” (1961). Retrospective Theses and Dissertations. Paper 2402
Haldane, J. S.. 1901. The Red Colour of Salted Meat. Journal of Hygiene 1: 115 – 122
Hoagland, R. 1914. Cloring matter of raw and cooked salted meats. Laboratory Inspector, Biochemie Division, Bureau of Animal Industry. Journal of Agricultural Research, Vol. Ill, No. 3 Dept. of Agriculture, Washington, D. C. Dec. 15, 1914.
Lemberg, R. and Legge, J. W.. 1949. Hematin Compounds and Bile Pigments. Interscience Publishers, Inc.
Soltanizadeh, N., Kadivar, M.. 2012. A new, simple method for the production of meat-curing pigment under optimised conditions using response surface methodology. Meat Science 92 (2012) 538–547 Elsevier Ltd.
http://cbs.umn.edu/academics/departments/bmbb/about/history/timeline
Images:
Image 1: Ralph Hoagland. Oakland Tribune, 5 July 1927