
Einleitung zu Speck & die Kunst zu Leben
Dies ist die Geschichte von Speck und Leben – unsere Geschichte. Sie reicht von den späten 1800er-Jahren, als große Fortschritte in der Kunst der Speckpökelung gemacht wurden, bis in die Gegenwart. Figuren, inspiriert von realen Personen, bewegen sich in einer Welt mit Steampunk-Flair, in der moderne Sprache und Technologie kunstvoll auf ein historisches Setting gelegt sind.
Es ist eine technologische und zugleich persönliche Reise – eine Auswahl von Briefen, die Kristi und ich während unserer Reise um die Welt auf der Suche nach dem Geheimnis des perfekten Bacons an unsere Kinder und unsere Familie schreiben, ergänzt durch Kapitel, die wir viele Jahre später hinzufügten, um die Geschichte zu vervollständigen und ein vollständiges Bild der Entwicklung der Pökelung zu geben. Dabei entdecken wir die Kunst des Lebens.
Wo auch immer sie begann – sie geht weiter. Das ist unsere Geschichte.
Einführung in dieses Kapitel
Dieses Kapitel führt in die Chemie des Pökelns ein. Es richtet sich an Wissenschaftler.
Das Pökelmolekül
Dieses Kapitel soll Ihnen genügend Hintergrundwissen vermitteln, um die Grundlagen der Pökelung zu verstehen und einige ihrer komplexeren Aspekte zu würdigen. Es ist nicht als naturwissenschaftliches Lehrbuch gedacht, daher erlaube ich mir, die Inhalte vereinfacht darzustellen. Ich weise zum Beispiel nicht immer ausdrücklich darauf hin, wenn ich bei der einfachen Schreibweise von Nitrit als NOO über eine ionische Verbindung spreche. Außerdem habe ich Abschnitte mit dem Titel „Möchten Sie mehr wissen?“ eingefügt – für Leserinnen und Leser mit chemischem Hintergrund oder vertieftem Interesse. Lassen Sie mich Sie durch diese Konzepte führen – eine atemberaubende Geschichte erwartet Sie! Versuchen Sie nicht, sich alle neuen Begriffe zu merken oder sofort alle Zusammenhänge zu verstehen. Lesen Sie einfach von Anfang bis Ende, auch wenn es stellenweise anspruchsvoll wird. Ich versichere Ihnen: Sie werden sich viel mehr merken, als Sie jetzt glauben.
Was ist Fleischpökelung?
Pökelung ist die Kunst, frisches Fleisch durch einen Verwandlungsprozess zu führen, der es widerstandsfähig gegenüber dem Verderb macht – selbst ohne Kühlung. Dabei entfalten sich Farbe und Geschmack auf eine Weise, die rohem Fleisch fremd ist. Während sich der Prozess vollzieht, treten zwei sensorische Veränderungen deutlich hervor. Zuerst entwickelt sich ein herzhafter, komplexer Geschmack, zugleich erdig und lebendig. Danach verändert sich die Farbe: Das Fleisch nimmt seine charakteristische rosige Tönung an.
Doch jenseits von Geschmack und Aussehen geschieht noch etwas viel Wichtigeres. Das Fleisch wird sicherer. Mikrobielle Bedrohungen werden zurückgedrängt, der Verderb verlangsamt sich. Im Zentrum dieser stillen Verwandlung steht ein einziger, unsichtbarer Akteur: Stickstoffmonoxid (NO). Dieses winzige Molekül trifft auf das Eisen in den Hämproteinen und löst eine Kettenreaktion aus, die Farbe stabilisiert, den Geschmack vertieft und das Fleisch vor Zersetzung schützt (Pearson & Gillett, 1996; Pegg & Shahidi, 2000). Lassen Sie uns die faszinierenden Mechanismen dahinter genauer betrachten.
Proteine, Hämproteine und Sauerstoff
Muskulatur besteht aus Proteinen. Unter den vielen Proteinarten im Muskel sind für uns insbesondere die Hämproteine von Bedeutung. Dabei handelt es sich um Proteine, die eine sogenannte Hämgruppe tragen – ein nicht-proteinogenes Molekül, das spezielle chemische Reaktionen ermöglicht, wie etwa die Bindung von Sauerstoff, den Elektronentransfer oder Redoxprozesse (Keilin, 1966; Nicholls und Ferguson-Miller, 2013).
Während der Pökelung bindet Stickstoffmonoxid an das Eisenatom im Protein Myoglobin und bildet Nitrosomyoglobin. Diese Reaktion erzeugt die stabile rosa Farbe gepökelten Fleisches und schützt es zugleich vor Oxidation und mikrobiellem Verderb (Pegg und Shahidi, 2000; Honikel, 2008). Myoglobin selbst ist ein kugelförmiges Protein, das im Muskelgewebe vorkommt. Es spielt eine zentrale Rolle bei der Speicherung und Freisetzung von Sauerstoff in Muskelzellen. Im Zentrum des Moleküls befindet sich eine Hämgruppe mit einem Eisenatom im zweiwertigen Zustand (Fe²⁺), das Sauerstoff binden kann und frischem Fleisch seine rote Farbe verleiht.
Die Struktur von Myoglobin ähnelt einer der vier Untereinheiten des Hämoglobins – jenem sauerstofftragenden Protein, das in den roten Blutkörperchen vorkommt. Während Myoglobin in den Muskelzellen wirkt, zirkuliert Hämoglobin im Blut. Beide Proteine enthalten Hämgruppen mit eisenhaltigen Zentren, die Sauerstoff binden können. Diese gemeinsame Hämstruktur verleiht sowohl Blut als auch Fleisch ihre rote Farbe und ihre Fähigkeit zur Sauerstoffbindung.
Häm, in diesem Zusammenhang, bezieht sich speziell auf die eisenhaltige Komponente des Proteins. Im Zentrum befindet sich ein Eisenatom, das in einen Porphyrinring eingebettet ist.
Die biologische Nützlichkeit von Eisen liegt in seiner Fähigkeit, zwischen zwei Zuständen zu wechseln: Fe²⁺ und Fe³⁺. Diese Redox-Flexibilität erlaubt es dem Eisen, ein einzelnes Elektron aufzunehmen oder abzugeben, was es ideal für Elektronentransportsysteme macht. Im Fe²⁺-Zustand (zweiwertig, ferrous) kann Eisen Sauerstoff binden, wodurch Hämoglobin und Myoglobin diesen speichern oder transportieren können. Wird Eisen zu Fe³⁺ (dreiwertig, ferric) oxidiert, kann es keinen Sauerstoff mehr binden. Dieses reversible Umschalten ermöglicht es dem Körper, Sauerstoff genau dort aufzunehmen und freizusetzen, wo er gebraucht wird. Der Übergang zwischen diesen beiden Zuständen kann durch die Redoxbedingungen der Umgebung, nahegelegene Moleküle, die Bindung oder Freisetzung von Sauerstoff selbst oder durch Stickstoffmonoxid unter Pökelbedingungen ausgelöst werden.
Dieser einfache Wechsel zwischen Elektronenzuständen erlaubt es dem Eisen, als molekulares Transportmittel in verschiedenen essenziellen Systemen zu dienen. Im Hämoglobin bindet und gibt Eisen Sauerstoff ab, um den Transport im Blutkreislauf zu ermöglichen. In den Mitochondrien sind eisenhaltige Proteine Teil der Elektronentransportkette und leiten Elektronen durch eine Reihe von Trägern weiter, um zelluläre Energie zu erzeugen.
Myoglobin und cytochromähnliche Proteine, die am Pökelprozess beteiligt sind, teilen sich dieses eisenhaltige Zentrum und dieselbe Redox-Flexibilität. Cytochrome sind Hämproteine, die in den Mitochondrien der Muskelzellen vorkommen. Obwohl sie die Fleischfarbe nicht beeinflussen, sind sie entscheidend für den Elektronentransfer im Prozess der ATP-Bildung – der Energieeinheit der Zelle. Diese cytochromähnlichen Proteine tragen Eisen in ihrem Zentrum und durchlaufen Redoxzyklen ähnlich wie Myoglobin. Ihr Einfluss auf den Pökelvorgang ist gering, dennoch gehören sie zur biochemischen Gesamtlandschaft des Muskelgewebes.
Sauerstoff ist für die Energiegewinnung unentbehrlich, da er als letzter Elektronenakzeptor in der mitochondrialen Elektronentransportkette dient. Dort verbindet er sich mit Elektronen und Protonen zu Wasser – eine Reaktion, bei der Energie freigesetzt wird, die zur Synthese von ATP genutzt wird.
Die zelluläre Atmung erzeugt ATP in drei Hauptphasen. Zunächst spaltet die Glykolyse im Zytosol Glukose und setzt dabei eine geringe Menge ATP sowie Elektronenträger frei. Danach erzeugt der Krebszyklus (auch Zitronensäurezyklus oder TCA-Zyklus) in den Mitochondrien große Mengen an NADH und FADH₂ – Moleküle, die Elektronen transportieren. Schließlich nutzt die Elektronentransportkette diese Elektronen, um einen Protonengradienten zu erzeugen, der wiederum durch die ATP-Synthase zur Produktion von ATP genutzt wird.
Der letzte Schritt, bei dem Sauerstoff Elektronen aufnimmt, wird durch Cytochrom-c-Oxidase katalysiert, ein Hämprotein. Stickstoffmonoxid kann sich vorübergehend an das Häm- oder Kupferzentrum dieses Enzyms binden und dadurch die Atmung unter bestimmten Bedingungen wie Sauerstoffmangel oder Entzündungen verlangsamen. Auf diese Weise nutzt der Körper NO, um die Energieproduktion zu regulieren und oxidativen Stress zu vermeiden.
Obwohl Myoglobin nicht direkt an der ATP-Synthese beteiligt ist, befindet es sich sehr nahe an den Mitochondrien in Muskelzellen und fungiert als kurzfristiger Sauerstoffspeicher, der O₂ dort freisetzt, wo und wann er benötigt wird. Die Nähe des Myoglobins zu den Mitochondrien ermöglicht eine schnelle Diffusion und eine effiziente Sauerstoffnutzung bei hoher Aktivität.
Der Übergang zwischen Fe²⁺ und Fe³⁺ im Eisen erfolgt im Mikrosekundenbereich und geschieht kontinuierlich während des gesamten Lebens. Diese Kontrolle des Redoxzustandes ist grundlegend, um den Sauerstofffluss und eine stetige Energieproduktion aufrechtzuerhalten.
Die Energieausbeute aus der zellulären Atmung ist gewaltig. Ein einzelnes Glukosemolekül liefert etwa 36 bis 38 ATP-Moleküle. Hochgerechnet erzeugt und nutzt der menschliche Körper in Ruhe etwa 100 Watt Energie – vergleichbar mit einer normalen Glühbirne – und das mit bemerkenswerter Effizienz und Selbstregulierung. Er übertrifft menschengemachte Systeme bei Weitem in Präzision, Reaktionsfähigkeit und Reparaturfähigkeit.
Stickstoffmonoxid bewirkt mehr als nur die Stabilisierung der Fleischfarbe. Noch bevor es sich an das Häm-Eisen bindet, besitzt freies NO antimikrobielle Eigenschaften, insbesondere gegen anaerobe Krankheitserreger wie Clostridium botulinum. Einmal gebunden, verdrängt NO den Sauerstoff aus der Hämgruppe und stoppt so oxidative Reaktionen, die sonst Farbe, Geschmack und Fett abbauen würden. Das Pökeln erreicht also auf chemischem Weg, was Vakuumverpackung physikalisch leistet: Es neutralisiert Sauerstoff von innen heraus. Pökeln bietet sowohl antimikrobielle als auch antioxidative Schutzwirkung und gehört damit zu den wirkungsvollsten bekannten Konservierungsmethoden.
Stickstoffmonoxid ist unverzichtbar für die Funktion des Körpers und für das Pökeln von Fleisch
Die Fähigkeit von Eisen, Elektronen aufzunehmen und abzugeben, erlaubt es, den Energie- und Sauerstoffverbrauch im Körper zu regulieren. Eines der schädlichsten Moleküle, das in unseren Zellen entsteht, ist nicht – wie oft fälschlich behauptet – Nitrit, sondern Superoxid. Diese reaktive Sauerstoffspezies, O₂•⁻, tritt im Alter vermehrt auf. Um dies zu verstehen, muss man die Rolle von Sauerstoff im Energiestoffwechsel betrachten.
ATP, Adenosintriphosphat, ist das primäre Energiemolekül der Zelle. Es wird hauptsächlich in den Mitochondrien durch Zellatmung gebildet, bei der Elektronen von Nährstoffen auf Sauerstoff übertragen werden. Wenn dieses System reibungslos funktioniert, wird Sauerstoff vollständig zu Wasser reduziert und die Energie sicher als ATP gespeichert. Mit zunehmendem Alter verschlechtert sich jedoch dieses System. Statt Elektronen paarweise aufzunehmen, nimmt Sauerstoff sie einzeln auf. Diese unvollständige Reduktion führt zur Bildung von reaktiven Sauerstoffspezies wie Superoxid, Wasserstoffperoxid und Hydroxylradikalen. Diese Moleküle schädigen Proteine, Fette und DNA.
Obwohl Superoxid gefährlich ist, verfügt der Körper über Mechanismen, es zu kontrollieren. Einer der wichtigsten ist Stickstoffmonoxid. Dieses kleine Radikalmolekül mit einem einzelnen ungepaarten Elektron spielt eine Schutzrolle, indem es reguliert, wie Elektronen übertragen werden. NO konkurriert mit Sauerstoff um die Bindung an Eisen in Enzymen wie Cytochrom-c-Oxidase, dem Endenzym der Elektronentransportkette. Durch die Glättung des Elektronenflusses hilft NO, die unvollständigen Sauerstoffreduktionen zu verhindern, die Radikale erzeugen.
Gerade im Alter ist das besonders wichtig, da zwei Veränderungen gleichzeitig auftreten. Erstens nimmt die Bildung von Superoxid zu. Zweitens sinkt die Fähigkeit des Körpers, Stickstoffmonoxid zu bilden. NO entsteht im Körper auf zwei Hauptwegen. Zum einen wirkt das Enzym Stickstoffmonoxidsynthase auf die Aminosäure L-Arginin und bildet daraus NO. Zum anderen nehmen wir Nitrat und Nitrit über die Nahrung auf, die der Körper auf mehreren Wegen – insbesondere unter Sauerstoffmangel – in NO umwandeln kann. Dies macht die Zufuhr von Nitrit über die Nahrung nicht nur sicher, sondern im Alter zunehmend wichtig.
In diesem einen Absatz wurden auch die beiden Wege eingeführt, auf denen das Pökeln funktioniert. Erstens: aus dem Fleisch selbst, wenn Proteine abgebaut und L-Arginin zu Stickstoffmonoxid umgesetzt wird. Zweitens: durch Zugabe von Nitrat oder Nitrit bei der Verarbeitung, wobei Nitrit im Fleisch chemisch zu NO umgewandelt wird. Diese doppelte Entstehung bildet das Herzstück des Pökelvorgangs und ist Thema dieses Kapitels. Doch bevor wir dorthin zurückkehren, muss das Wirken von Stickstoffmonoxid allgemein verstanden werden.
Während NO seine Hauptfunktion durch Bindung an Häm-Eisen in Enzymen wie der Cytochrom-c-Oxidase erfüllt, wirkt es auch als Verteidiger gegen Radikale. Zwar sucht NO Superoxid nicht aktiv auf wie ein bewegliches Molekül, doch es reagiert mit Superoxid zu Peroxynitrit. Peroxynitrit ist ebenfalls reaktiv, löst jedoch weitaus seltener jene schädlichen Kettenreaktionen aus, die freies Superoxid verursacht. Auf diese Weise unterbricht NO die Entstehung von Radikalkaskaden und begrenzt Zellschäden.
Es sollte nun klar sein, dass Sauerstoff – obwohl lebensnotwendig – auch gefährlich sein kann. Wenn er bei der Atmung nicht vollständig reduziert wird, entstehen Radikale, die Zellen schädigen. Stickstoffmonoxid verhindert dies, indem es an Eisen in Schlüsselenzymen bindet, den Elektronenfluss reguliert und somit die Bildung dieser Radikale einschränkt. Im Blutkreislauf kann NO auch an Hämoglobin binden. Zwar reguliert es dadurch nicht direkt die mitochondriale Atmung, doch beeinflusst es, wie viel NO im Blut verfügbar bleibt. Je nach Sauerstoffgehalt im Gewebe kann Hämoglobin NO speichern oder freisetzen. So sorgt es dafür, dass sich Blutgefäße dort erweitern, wo es nötig ist, und der Sauerstoff effizient verteilt wird. Auf diese Weise schützt NO sowohl die Zellfunktion als auch das Gefäßsystem.
Dieses Prinzip gilt auch für gepökeltes Fleisch. So wie NO an das Eisen in mitochondrialen Enzymen bindet, bindet es auch an das Eisen im Myoglobin. Dadurch wird verhindert, dass Sauerstoff oxidative Prozesse auslöst, die Farbe und Geschmack des Fleisches verschlechtern würden. In der Biologie wie beim Pökeln ist die Chemie dieselbe: NO besetzt das Eisen im Myoglobin und blockiert den durch Sauerstoff angetriebenen Redoxkreislauf. So wird die Bildung von Metmyoglobin – dem braunen Pigment, das auf Verderb hinweist – gestoppt und Fette vor dem Ranzigwerden geschützt. Ob in einer lebenden Zelle oder in einem Stück Schinken – Stickstoffmonoxid schützt, indem es die mächtige, potenziell zerstörerische Chemie des Sauerstoffs kontrolliert.
Kurz gefasst
Wie NO die Bildung von Radikalen verhindert
Reaktive Sauerstoffspezies (ROS) entstehen, wenn Sauerstoff nur teilweise reduziert wird – also Elektronen einzeln aufnimmt. Stickstoffmonoxid (NO) begrenzt dies durch folgende Mechanismen:
- Es konkurriert mit O₂ an Metallzentren (Häm und Nicht-Häm) und verhindert so eine vorzeitige Sauerstoffreduktion.
- Es interagiert mit metallhaltigen Enzymen und verlangsamt den Elektronenverlust.
- Es reagiert mit Radikalen (wobei unter Umständen reaktive Stickstoffspezies wie Peroxynitrit, ONOO⁻, entstehen – auch diese sind reaktiv, daher ist der Kontext entscheidend).
Im Fleisch liegt die farbstabilisierende und antioxidative Wirkung von NO sowohl in seiner Bindung an das Pigment als auch in seiner Fähigkeit zur Radikalbindung begründet (Beckman & Koppenol, 1996; Gray, Gomaa & Buckley, 1996; Pegg & Shahidi, 2000).
Wie entsteht Stickstoffmonoxid?
Stickstoffmonoxid, oder NO, wird im menschlichen Körper über zwei Hauptwege gebildet. Der erste ist der sogenannte L-Arginin-Weg, bei dem spezialisierte Enzyme – die sogenannten Stickstoffmonoxidsynthasen – die Aminosäure L-Arginin in Stickstoffmonoxid und L-Citrullin umwandeln. Dieser Weg ist für den größten Teil des natürlich im Körper gebildeten NO verantwortlich, insbesondere in Blutgefäßen, im Nervensystem und in Immunzellen. Der zweite Weg ist diätetischer Natur. Dabei werden Nitrat und Nitrit, die in der Nahrung vorkommen, vor allem unter Bedingungen mit wenig Sauerstoff oder niedrigem pH-Wert in Stickstoffmonoxid umgewandelt. Schätzungen variieren, aber allgemein geht man davon aus, dass etwa 60–70 % des Stickstoffmonoxids durch körpereigene Enzymsysteme gebildet werden, während die restlichen 30–40 % über die Nahrung zugeführt werden.
Zu den Nahrungsquellen gehören nitratreiche Gemüsesorten wie Spinat, Rote Bete, Salat und Sellerie. Eine weitere wichtige Quelle ist gepökeltes Fleisch. Produkte wie Speck oder Schinken enthalten kleine, gesetzlich regulierte Mengen an Restnitrit, das im Körper, insbesondere im sauren Milieu des Magens, als Substrat für die Bildung von NO dienen kann. Dieses Nitrit ist keineswegs schädlich – im Gegenteil: Es kann einen wichtigen Beitrag zur Aufrechterhaltung eines gesunden NO-Spiegels leisten, insbesondere im Alter, wenn die körpereigene Produktion abnimmt.
Auch beim Pökeln von Fleisch entsteht Stickstoffmonoxid durch zwei Hauptmechanismen.
Der erste Mechanismus ist mikrobiell und basiert auf dem L-Arginin-Weg. Während das Fleisch reift, werden Muskelproteine abgebaut und L-Arginin wird im Gewebe verfügbarer. Unter geeigneten mikrobiellen Bedingungen wird dieses L-Arginin in NO umgewandelt. Dieser natürliche Prozess ist die Grundlage vieler sogenannter „nitritfreier“ oder „natürlich gepökelter“ Fleischprodukte. Viele Menschen, die versuchen, Fleisch ohne Zusatz von Nitrit zu pökeln – oft aus Angst vor angeblichen Gesundheitsrisiken – sind überrascht, wenn sich nach einigen Tagen eine rosa Färbung zeigt, zumindest stellenweise. Dies ist das Ergebnis einer langsamen Umwandlung von L-Arginin zu NO durch Bakterien, die eine partielle Pökelreaktion auslöst.
Der zweite Mechanismus ist der klassische Weg von Nitrat über Nitrit zu Stickstoffmonoxid. Traditionell wurde dem Fleisch Kaliumnitrat – auch bekannt als Salpeter – zugesetzt. Bakterien reduzierten dann das Nitrat zu Nitrit, welches anschließend chemisch zu NO reduziert wurde. Dieses NO bindet sich an das Häm-Eisen des Fleischpigments und verleiht dem Fleisch die typische Pökelfarbe und das charakteristische Aroma. In der heutigen industriellen Praxis wird der erste bakterielle Schritt oft übersprungen, indem man direkt Natriumnitrit einsetzt. Dies sorgt für eine schnellere und kontrollierbare Reaktion. Auch bei der Verwendung pflanzlicher Pökelmittel wie Sellerie- oder Rote-Bete-Pulver ist die chemische Abfolge dieselbe: Das enthaltene Nitrat oder Nitrit wird im Fleisch letztlich in NO umgewandelt.
Es ist wichtig zu verstehen, dass sowohl die Nitrat-basierte als auch die L-Arginin-basierte Pökelung von mikrobieller Aktivität abhängen. Jahrzehntelang wurde übersehen, dass L-Arginin als Stickstoffquelle für das Pökeln dienen könnte. Das liegt unter anderem daran, dass Bakterien bei Vorhandensein von Nitrat dieses bevorzugt nutzen. Die Rolle von L-Arginin wird erst dann sichtbar, wenn kein Nitrat vorhanden ist. Erste Vermutungen, dass L-Arginin bei langzeitgereiften Produkten wie traditionellem Rohschinken eine Rolle spielen könnte, wurden schließlich bestätigt. In den 1990er Jahren konnten Forscher nachweisen, dass Bakterien diese Umwandlung von L-Arginin zu NO vermitteln können. Anfangs schien dies nur für lange gereifte Produkte zu gelten. In den letzten Jahren jedoch wurden kommerzielle, fermentationsbasierte Systeme entwickelt, mit denen Fleisch allein durch bakterielle Aktivität – also ohne Zugabe von Nitrit oder Nitrat – schnell gepökelt werden kann. Dabei entsteht Stickstoffmonoxid direkt durch Fermentation und die Pökelgeschwindigkeit entspricht jener bei Verwendung von Natriumnitrit.
Dennoch: Ohne die Abkürzung durch Nitrit verläuft die NO-Bildung langsamer und unberechenbarer. Nitrit bietet einen direkten chemischen Weg zur NO-Bildung, ohne dass eine langsame bakterielle oder enzymatische Umwandlung nötig wäre. Fehlt Nitrit, hängt der gesamte Prozess von der Verfügbarkeit geeigneter Bakterien, den notwendigen Enzymen sowie von optimalem pH-Wert, Temperatur und mikrobieller Balance ab. Das erklärt, warum sich das Nitritpökeln – ob durch zugesetztes Nitrat oder Nitrit – über Jahrhunderte hinweg als die zuverlässigste Methode etabliert hat, um Fleisch sicher zu konservieren, Geschmack und Farbe zu entwickeln und vor mikrobiellen Verderb zu schützen.
Damit ist der Grundstein für unsere Betrachtung von Pökelsystemen gelegt. Ich wusste schon vor vielen Jahren, dass Bakterien L-Arginin in NO umwandeln können. Doch es dauerte wesentlich länger zu verstehen, was nötig ist, um diesen Weg verlässlich in einem Pökelsystem zu nutzen. Es reicht nicht, einfach Salz hinzuzufügen und auf Farbe zu hoffen. Die Farbstabilität muss gezielt gesteuert werden. Das mikrobiologische Umfeld muss sicher sein. Biofilmbildung muss verhindert werden. Die Reaktionsgeschwindigkeit muss erhöht werden. All das geschieht nicht von selbst – es muss mit Fachwissen und Sorgfalt geplant und umgesetzt werden. Es gehört zu den anspruchsvollsten, aber auch lohnendsten Aufgaben in der Welt des Pökelns.
Kehren wir noch einmal zurück zu den beiden Hauptreaktionen, die wir gerade betrachtet haben. Es handelt sich dabei nicht nur um chemische Abläufe. Es sind zwei verschiedene Wege – zwei Denkkarten –, mit denen der Mensch gelernt hat, mikrobiellen und oxidativen Verderb zu kontrollieren und gleichzeitig Geschmack und Farbe im Fleisch hervorzubringen. Im Laufe dieses Kapitels werden wir immer wieder zu diesen beiden Reaktionswegen zurückkehren. Die Geschichte des Specks ist im Kern die Geschichte dieser beiden Wege – und davon, wie das Pökeln stets darauf abzielte, Stickstoffmonoxid so schnell und effektiv wie möglich zu erzeugen.
Wissenschaftler und Pökelmeister haben seit Langem nach Wegen gesucht, auf Nitrit in der Fleischverarbeitung zu verzichten. Meist geschah dies aus Sorge um mögliche Gesundheitsrisiken – Sorgen, die oft übertrieben und schlecht verstanden sind. Die bakterielle Fermentation von L-Arginin bietet einen Weg, dieses Ziel zu erreichen. Doch eines möchte ich von Anfang an unmissverständlich klarstellen: Stickstoffmonoxid, Nitrit und Nitrat sind wie drei eng verwandte Substanzen. Wo man eine findet, findet man auch die anderen. Ihre Reaktivität sorgt für ständigen Austausch. Selbst wenn man NO ausschließlich aus L-Arginin erzeugt, entstehen zwangsläufig auch Nitrit und Nitrat im Fleisch. Es ist chemisch unmöglich, gepökeltes Fleisch mit NO zu erzeugen, ohne dabei auch etwas Nitrit zu bilden. Das sollte jedoch nicht als Problem gesehen werden. Wie wir gezeigt haben, spielt Nitrit eine wichtige physiologische Rolle im menschlichen Körper. Es ist nicht nur sicher – es ist nützlich. Und je älter wir werden, desto wichtiger wird sein Beitrag.
Möchten Sie mehr erfahren?
Ein genauerer Blick auf die Nitrat–Nitrit–Stickstoffmonoxid-Kaskade im menschlichen Körper
Es ist seit Langem bekannt, dass der Nitrat–Nitrit–Stickstoffmonoxid-(NO)-Weg im menschlichen Körper funktioniert und durch die Aufnahme von Nitrat über die Nahrung eingeleitet wird. Dieser Weg ist keineswegs auf das Pökeln von Fleisch beschränkt. Er stellt ein zentrales physiologisches System dar, das die grundlegende Biochemie widerspiegelt, die das Leben aufrechterhält. Wie Weitzberg (2010) und andere gezeigt haben, ist der Nitrat–Nitrit–NO-Weg weit mehr als nur ein sekundärer Mechanismus zur NO-Bildung. Gemeinsam mit ihm hat Jon O. Lundberg (Karolinska-Institut, Schweden) bahnbrechende Studien zu den systemischen Effekten von Nitrat in der menschlichen Gesundheit verfasst. Nathan S. Bryan (University of Texas Health Science Center, Houston) hat intensiv die Umwandlungswege von Nitrat untersucht und deren Bedeutung für die Regulation des Herz-Kreislauf-Systems betont. Alan N. Schechter (National Institutes of Health, USA) war ein Pionier in der Erforschung von Nitrit als NO-Reservoir, insbesondere bei Sauerstoffmangel. Zhongxin Zhuge (University of California, San Francisco) trug wichtige mechanistische Erkenntnisse zur Nitrit-zu-NO-Umwandlung in Geweben bei. Mark Gladwin (University of Pittsburgh School of Medicine) wurde durch seine Forschung zur NO-Bildung aus Nitrit im Blut und Gefäßgewebe bekannt, vor allem unter hypoxischen Bedingungen. Gemeinsam zeichneten diese Wissenschaftler das Bild eines physiologischen Systems, das dem Pökelprozess ähnelt – und für das Leben selbst unverzichtbar ist.
Beim Menschen wird Nitrat (NO₃⁻) hauptsächlich über nitratreiche Nahrungsmittel aufgenommen, darunter Blattgemüse, Wurzelgemüse und gepökelte Fleischwaren. Nach der Aufnahme gelangt das Nitrat in den Blutkreislauf und wird in den Speicheldrüsen konzentriert. Dort wandeln Mundbakterien es in Nitrit (NO₂⁻) um, das geschluckt wird und im sauren Milieu des Magens landet. In Anwesenheit von Reduktionsmitteln wie Ascorbinsäure (Vitamin C) und Polyphenolen wird das Nitrit dann chemisch zu Stickstoffmonoxid (NO) reduziert – besonders unter sauerstoffarmen (hypoxischen) oder sauren Bedingungen.
Diese Forscher haben gezeigt, dass es sich dabei keineswegs nur um einen Notmechanismus handelt. Vielmehr funktioniert der Nitrat–Nitrit–NO-Weg als eigenständiges Regulationssystem. Verschiedene Studien belegen, dass er den klassischen L-Arginin–NO-Synthase-(NOS)-Pfad ergänzt, insbesondere dann, wenn dieser beeinträchtigt ist – etwa bei Hypoxie oder bei begrenzter Verfügbarkeit von L-Arginin. In solchen Fällen sichert der Nitrat–Nitrit-Weg die kontinuierliche NO-Bildung und schützt damit zentrale physiologische Prozesse.
Bemerkenswert ist, dass diese Abfolge – von Nitrat über Nitrit zu Stickstoffmonoxid – exakt derselben chemischen Reaktion entspricht, die auch beim traditionellen Pökeln mit Salpeter oder Natriumnitrit abläuft. So wie dieser Weg im Fleisch die Farbe stabilisiert und das mikrobielle Wachstum hemmt, reguliert er im menschlichen Körper den Gefäßtonus und schützt Gewebe vor oxidativem Stress.
Schätzungen zufolge stammen etwa 30 bis 40 % der gesamten NO-Produktion im menschlichen Körper aus dem Nitrat–Nitrit–NO-Weg, während die übrigen 60 bis 70 % durch NOS-Enzyme aus L-Arginin gebildet werden. Dieses doppelte System sorgt nicht nur für Redundanz bei der NO-Bildung, sondern erlaubt dem Körper auch eine präzise Feinsteuerung der NO-Konzentration in Abhängigkeit von physiologischen und umweltbedingten Bedingungen. Es ist ein eindrucksvolles Beispiel dafür, wie sich Lebensmittelbiochemie, Fleischwissenschaft und menschliche Gesundheit auf natürliche Weise überschneiden.
Können wir Stickstoff (Nitrat oder Nitrit) aus unserer Ernährung entfernen?
Wir alle wissen, wie wichtig Sauerstoff für unser tägliches Leben ist. Ohne ihn wäre Leben, wie wir es kennen, nicht möglich. Ein zweites Element, das ebenso lebensnotwendig ist wie Sauerstoff, ist Stickstoff. Woher kommt Stickstoff – und warum ist er so entscheidend für das Leben? Bevor wir uns wieder Nitrat und Nitrit in der Nahrung und der Pökelchemie zuwenden, lohnt es sich, kurz einen Schritt zurückzutreten und Stickstoff als solchen zu betrachten.
Die Bedeutung von Stickstoff
Stickstoff ist eines der grundlegenden Elemente des Lebens. Im Pflanzenreich wird Stickstoff hauptsächlich in Form von Nitrat oder Ammonium aus dem Boden aufgenommen und in Aminosäuren, Proteine und Nukleinsäuren eingebaut – Moleküle, die für Wachstum, Energieübertragung und Fortpflanzung unentbehrlich sind. Das satte Grün gesunder Pflanzen ist direkt mit der Stickstoffverfügbarkeit verknüpft, da Chlorophyll, das Pigment der Photosynthese, Stickstoff als zentrales Strukturelement enthält.
Wenn Tiere Pflanzen fressen, übernehmen sie diesen Stickstoff in organisch gebundener und biologisch verfügbarer Form. Der Nährwert von Weide- und Futtermitteln hängt in hohem Maße vom Stickstoffgehalt ab, der wiederum eng mit dem Proteingehalt korreliert. Zahlreiche frühe Experimente in der Tierphysiologie zeigten einen klaren Zusammenhang zwischen Stickstoffaufnahme und Überleben. Tiere, die mit stickstofffreier Nahrung gefüttert wurden, bauten rasch körperlich ab und starben ohne rechtzeitige Intervention an Stickstoffmangel. Im Gegensatz dazu entwickelten sich Tiere, die proteinreiche – also stickstoffreiche – Nahrung erhielten, gesund und kräftig. Diese Versuche bewiesen, dass Stickstoff nicht nur ein nützlicher Nährstoff ist, sondern ein absolut lebensnotwendiger.
Möchten Sie mehr erfahren?
Die Rolle von Stickstoff in Pflanzen
Stickstoff ist Bestandteil des grünen Pflanzenpigments namens Chlorophyll, das für die Photosynthese verantwortlich ist. Darüber hinaus fördert Stickstoff das schnelle Wachstum der Pflanze, steigert die Samen- und Fruchtbildung und verbessert die Qualität von Blattmasse und Futterpflanzen (Plant Nutrients and Lilies).
Dies ist insofern bedeutsam, als wir später sehen werden, dass Nitrat, Nitrit und Stickstoffmonoxid nicht nur Fleisch pökeln und die Gesundheit unseres Körpers unterstützen, sondern dass dieselbe Reaktion auch für die Ernährung von Pflanzen zentral ist. Der Pökelvorgang ist also keineswegs ein künstlicher oder fremder Prozess. Im Gegenteil – er ist lebenswichtig für sämtliche Aspekte tierischen und pflanzlichen Lebens. Und der Mensch gehört selbstverständlich zu dieser Gruppe von Lebewesen.
Stickstoff als Pflanzennahrung
Kalium (K) und Stickstoff (N) zählen zusammen mit Phosphor (P) zu den wichtigsten Nährstoffen für Pflanzen. Diese sogenannten Hauptnährstoffe sind im Boden häufig in unzureichender Menge vorhanden, weil Pflanzen sie beim Wachstum aufnehmen und somit den Boden auslaugen. Wie wir später sehen werden, erneuert die Natur den Stickstoffkreislauf auf vielfältige Weise, doch die moderne Landwirtschaft hat den Bedarf geschaffen, zusätzlich Stickstoff in den Boden einzubringen. Kalium, Stickstoff und Phosphor gehören zu den sogenannten Makronährstoffen. Die sogenannten Sekundärnährstoffe sind Kalzium (Ca), Magnesium (Mg) und Schwefel (S). Diese kommen im Boden meist in ausreichenden Mengen vor. Wenn Kalk auf sauren Böden ausgebracht wird, werden große Mengen Kalzium und Magnesium zugeführt. Schwefel entsteht in der Regel durch den Abbau organischer Substanz. Kalium wird in größeren Mengen aufgenommen als jeder andere Mineralstoff außer Stickstoff – in manchen Fällen sogar mehr als Kalzium. Kalium unterstützt den Aufbau von Proteinen, die Photosynthese, die Fruchtqualität und verringert das Auftreten von Pflanzenkrankheiten (Plant Nutrients and Lilies). Der historische Ursprung des Begriffs „Kalium“ liegt übrigens in der Asche von Pflanzen: Die Bezeichnung stammt von „Pottasche“ – also Pflanzenasche, die in einem Topf mit Wasser ausgelaugt wurde – und geht der Industriellen Revolution voraus.
Alle Proteine – also die Bausteine unserer Muskeln – enthalten Stickstoff. Unser Körper nutzt Stickstoffmonoxid auf viele verschiedene Weisen, um gesund zu bleiben. So sehr, dass Leben ohne Stickstoffmonoxid im Körper nicht möglich wäre. Die entscheidende Frage ist: Produziert der Körper genug NO selbst? Die Antwort lautet: Nein. Wir müssen das, was der Körper nicht selbst herstellen kann, über unsere Nahrung zuführen. Zu den Nahrungsmitteln, die Nitrat oder Nitrit enthalten, zählen:
Gemüse: Die bei weitem bedeutendste Quelle von Nitrat ist grünes Blattgemüse. Die Art und Weise, wie Nitrat in unserem Körper zu Stickstoffmonoxid wird, verläuft über die sogenannte Nitrat–Nitrit–NO-Kaskade. Diese Gemüsesorten enthalten auch Nitrit, das ebenfalls über verschiedene chemische Zwischenschritte in NO umgewandelt wird: Nitrit → verschiedene Reaktionen → NO.
Wasser: Grundwasser enthält häufig Nitrat oder Nitrit aus tierischen und menschlichen Abfällen sowie aus Düngemitteln in der Umgebung. Die chemische Reaktionskette ist dieselbe wie beim Gemüse: Nitrat → Nitrit → NO.
Gepökeltes Fleisch: Nitratverbindungen kommen weltweit natürlich vor. Kaliumsalpeter (KNO₃) ist ein bekanntes Beispiel. Nitritsalze sind hingegen hergestellte Verbindungen aus Natrium und Nitrit. Salpeter (also Kalium- oder Natriumnitrat) wird auch heute noch beim Pökeln verwendet. Wenn wir gepökeltes Fleisch essen, nehmen wir Nitrat oder Nitrit auf, das im Körper über die Kaskade Nitrat → Nitrit → NO oder direkt über Nitrit → NO in Stickstoffmonoxid umgewandelt wird. Gepökeltes Fleisch ist jedoch mengenmäßig die kleinste und unbedeutendste Quelle von Nitrat und Nitrit in unserer Ernährung.
Der Weg vom Nitrat zum Stickstoffmonoxid ist zentral. Lassen Sie uns das an einem Beispiel verdeutlichen – dem Salpeter. Salpeter besteht aus einem Stickstoffatom und drei Sauerstoffatomen – um es anschaulich zu machen, schreiben wir es als NOOO, wobei wir den metallischen Anteil der Verbindung (Kalium oder Natrium) ignorieren. Das Nitrat bildet mit Metallen wie Natrium, Kalzium oder Kalium stabile Salze wie Natriumnitrat, Kalziumnitrat oder Kaliumsalz (Salpeter). Für die Fleischverarbeitung spielt nur das Natrium noch eine zusätzliche Rolle – darauf gehen wir später ein. Für den Moment genügt es, sich auf den Nitratanteil zu konzentrieren.
Wenn Nitrat an eines dieser Metalle gebunden ist, bildet es ein sehr stabiles Salz, das seinen zusätzlichen Sauerstoff nicht leicht abgibt. Wir schreiben es als NOOO – tatsächlich ist es NO₃⁻. Diese stabile Verbindung kann durch Bakterien ein Sauerstoffatom verlieren, welches sie für ihren Stoffwechsel verwenden. NOOO wird dadurch zu NOO, also Nitrit (NO₂⁻). Im Gegensatz zum Nitrat ist Nitrit eine instabile Verbindung, die leicht weiter reagiert und in andere reaktive Stickstoffverbindungen wie NO übergeht. Wenn NOO ein weiteres Sauerstoffatom verliert, entsteht NO, Stickstoffmonoxid. Dieser Schritt ist chemischer Natur und nicht mikrobiell. Dabei entsteht NO meist über weitere Zwischenformen.
Die alten Pökelmethoden begannen mit Nitrat, das erst zu Nitrit und dann zu NO wurde. So wurde Fleisch konserviert, bevor es weltweit Zugang zu Natriumnitrit gab – also vor dem Ersten Weltkrieg. Viele handwerklich arbeitende Pökler bevorzugen bis heute den Start mit Nitrat, da die bakterielle Umwandlung zur Geschmacksentwicklung beiträgt. Wird direkt mit Nitrit gearbeitet, fällt dieser Prozess weg. Seit dem Zweiten Weltkrieg wurde es üblich, die Umwandlung von Nitrat zu Nitrit zu überspringen, da sie zeitaufwendig ist und zu inkonsistenten Ergebnissen führen kann. Heutige industrielle Methoden beginnen die Reaktion meist direkt mit Natriumnitrit.
Ob es nun der Weg Nitrat → Nitrit → NO oder Nitrit → NO ist – in beiden Fällen wird jeweils ein Sauerstoffatom entfernt. Der umgekehrte Weg ist jedoch ebenfalls möglich: Stickstoffmonoxid kann ein Sauerstoffatom aufnehmen und so zu NOO (Nitrit) werden, das wiederum zu NOOO (Nitrat) wird. Wo immer man also Nitrat, Nitrit oder NO findet, wird man mit hoher Wahrscheinlichkeit auch die jeweils anderen vorfinden. Diese reversiblen Reaktionen erklären, warum man in gepökeltem Fleisch meist Spuren aller drei Verbindungen findet – selbst wenn man nur eine davon zugesetzt hat.
Möchten Sie mehr erfahren?
Ionenverbindungen
Es ist leicht zu erkennen, dass die „3“ in NO₃⁻ (Nitrat) bedeutet, dass ein Stickstoffatom mit drei Sauerstoffatomen verbunden ist – aber was bedeutet das Minuszeichen? Der Stickstoff zusammen mit den drei Sauerstoffen bildet eine einzige, gemeinsam geladene Einheit. Die Nettoladung dieses „Pakets“ ist negativ – daher das „⁻“. Eine solche geladene Einheit bezeichnet man als Ion (wenn es sich um ein einzelnes Atom handelt) oder als Bestandteil einer Ionenverbindung (wenn mehrere Atome verbunden sind und zusammen eine Nettoladung tragen). Eine Verbindung ist ganz allgemein eine Gruppierung von zwei oder mehr chemischen Elementen (Atkins & de Paula, 2014; Housecroft & Sharpe, 2012).
Ionenverbindungen werden durch elektrostatische Kräfte zwischen entgegengesetzt geladenen Ionen zusammengehalten. Ein Kation ist positiv geladen, ein Anion negativ. Treffen sie im richtigen Mengenverhältnis aufeinander, ist das Gesamtsystem elektrisch neutral. Ein bekanntes Beispiel ist Kochsalz: NaCl, bestehend aus Na⁺ (Kation) und Cl⁻ (Anion).
Stickstoff und Sauerstoff bilden eine Gruppe sogenannter Oxo-Anionen, die für das Pökeln von großer Bedeutung sind – vor allem Nitrat (NO₃⁻) und Nitrit (NO₂⁻). So wie „Kochsalz“ die gebräuchliche Bezeichnung für Natriumchlorid ist, ist „Salpeter“ der gebräuchliche Name für Kaliumnitrat (KNO₃). Das Nitration (NO₃⁻) verbindet sich mit einem Metall-Kation (z. B. K⁺, Na⁺, Ca²⁺), um Salze wie KNO₃, NaNO₃ oder Ca(NO₃)₂ zu bilden. In Wasser kann man sich diese Salze als Kombination aus Salpetersäure (HNO₃) und einer Base wie Kaliumhydroxid (KOH) vorstellen, wobei ein kristallines Salz wie [K⁺][NO₃⁻] (kompakt: KNO₃) entsteht (Housecroft & Sharpe, 2012).
Ebenso bildet Nitrit (NO₂⁻) Salze mit denselben Metallen:
Nitratsalze
NOOO (Nitrat) + K (Kalium) → KNO₃ (Kaliumnitrat)
NOOO (Nitrat) + Na (Natrium) → NaNO₃ (Natriumnitrat)
NOOO (Nitrat) + Ca (Kalzium) → Ca(NO₃)₂ (Kalziumnitrat)
Nitritsalze
NOO (Nitrit) + K (Kalium) → KNO₂ (Kaliumnitrit)
NOO (Nitrit) + Na (Natrium) → NaNO₂ (Natriumnitrit)
NOO (Nitrit) + Ca (Kalzium) → Ca(NO₂)₂ (Kalziumnitrit)
In unseren Diskussionen zum Fleischpökeln wird das Metallion (K, Na, Ca) meist ignoriert, sobald das Salz in Lösung gegangen ist, weil die eigentliche Chemie durch die reaktiven Stickstoffverbindungen (Nitrat, Nitrit, Stickstoffmonoxid) gesteuert wird – also genau jene Verbindungen, die für Farbe, Geschmack und Haltbarkeit entscheidend sind (Pegg & Shahidi, 2000; Honikel, 2008).
Reduktion vs. Oxidation (in der einfachen Sauerstoff-Zählweise, wie sie hier verwendet wird)
Im Zusammenhang mit Stickstoffverbindungen lässt sich Reduktion und Oxidation auf einfache Weise über die Anzahl der Sauerstoffatome erklären. Reduktion bedeutet den Verlust von Sauerstoffatomen, während Oxidation den Zugewinn von Sauerstoffatomen beschreibt. Dieses vereinfachte Modell hilft dabei, die Umwandlungen zwischen Nitrat, Nitrit und Stickstoffmonoxid nachzuvollziehen.
Wenn Nitrat (vereinfacht als NOOO oder chemisch als NO₃⁻ dargestellt) ein Sauerstoffatom verliert, entsteht Nitrit (NOO oder NO₂⁻). Verliert Nitrit ein weiteres Sauerstoffatom, entsteht Stickstoffmonoxid (NO). Diese Abfolge steht im Zentrum sowohl der biologischen NO-Bildung als auch der traditionellen Fleischpökelung.
Umgekehrt beschreibt Oxidation den entgegengesetzten Prozess: Aus stärker reduzierten Stickstoffverbindungen wie Ammoniak (NH₃ oder NH₄⁺) können Sauerstoffatome – entweder biologisch oder chemisch – hinzugefügt werden, um Stickstoffmonoxid (NO) zu erzeugen oder weiter zu Nitrit (NO₂⁻) und Nitrat (NO₃⁻) aufzusteigen. Welcher Weg genommen wird, hängt von den jeweiligen Umweltbedingungen und den beteiligten Enzymen ab (Hanrahan, 2005; Lundberg, Weitzberg & Gladwin, 2008).
Diese Umwandlungen sind nicht nur theoretischer Natur – sie sind grundlegend für lebenswichtige Prozesse im menschlichen Körper sowie für bewährte Verfahren der Lebensmittelkonservierung. Auf die Geschichte der Umwandlung von Ammoniak zu Stickstoffmonoxid – insbesondere durch mikrobielle Nitrifikation – werden wir später noch zurückkommen, wenn wir die historische Bedeutung von Salmiak (sal ammoniac) und verwandten Pökelverfahren betrachten.
Reduktion (Verlust von Sauerstoffatomen)
NOOO → NOO → NO
Nitrat (NOOO / NO₃⁻) verliert ein O → Nitrit (NOO / NO₂⁻)
Nitrit verliert ein O → Stickstoffmonoxid (NO)
Oxidation (Zugewinn von Sauerstoffatomen)
Aus reduzierten Stickstoffverbindungen wie Ammoniak (NH₃ / NH₄⁺) können durch Hinzufügen von Sauerstoff chemisch oder biologisch NO, NO₂⁻ oder NO₃⁻ entstehen – je nach Reaktionsweg und Umweltbedingungen (Hanrahan, 2005; Lundberg, Weitzberg & Gladwin, 2008).
Wir werden auf die mikrobielle Umwandlung von Ammoniak zu NO zurückkommen, wenn wir Salmiak und verwandte historische Pökelprozesse näher untersuchen.
Die allgegenwärtige Präsenz von Stickstoff
Atmosphärischer Stickstoff (N₂) wird ständig in reaktive Stickstoffverbindungen umgewandelt, die in einem unaufhörlichen Kreislauf durch Pflanzen, Tiere, Wasser, Boden und Atmosphäre zirkulieren. Otto et al. (2010) schätzen, dass etwa 1,4 Milliarden Blitzeinschläge pro Jahr rund 8,6 Milliarden Tonnen NOx erzeugen – eine Gruppe aus Stickstoffoxiden. Das „x“ in NOx steht dabei für die variable Anzahl an Sauerstoffatomen: NO (x = 1), NO₂ (x = 2), NO₃⁻ (x = 3) usw.
Diese natürlichen Mengen übersteigen alles, was die Pökelindustrie je freisetzen könnte, bei Weitem. Daraus ergibt sich ein deutliches Argument gegen die Vorstellung, dass Nitrat, Nitrit oder NO „künstliche“ oder „industrielle“ Chemikalien seien, die dem Fleisch aufgezwungen werden. Diese Verbindungen sind allgegenwärtig, sie zirkulieren beständig und sind essenziell für das Leben (Hanrahan, 2005; Lundberg et al., 2008; Otto et al., 2010).
Nitrit ist – ebenso wie NO selbst – hochreaktiv und daher in der Umwelt seltener und kurzlebiger als Nitrat, das sehr stabil ist. Während heute der Großteil des verzehrten Natriumnitrits als industriell hergestelltes Salz in Umlauf kommt, ist Nitrat in der Natur reichlich vorhanden – besonders in Gemüse. NO dagegen ist flüchtig und wird rasch in andere Stickstoffverbindungen umgewandelt (Weitzberg & Lundberg, 2010; Honikel, 2008).
Möchten Sie mehr erfahren?
Die allgegenwärtige Präsenz von Stickstoff
„Stickstoff ist ein essentielles Element für alle Lebensformen und der strukturelle Baustein von Aminosäuren, aus denen tierisches und menschliches Gewebe, Enzyme und viele Hormone bestehen. Für das Pflanzenwachstum ist verfügbarer (fixierter) Stickstoff in natürlichen Systemen meist der begrenzende Nährstoff. Die Stickstoffchemie und ihr Kreislauf in der globalen Umwelt sind aufgrund der Vielzahl möglicher Oxidationsstufen äußerst komplex. Stickstoff besitzt fünf Valenzelektronen und kommt in Oxidationsstufen von −3 bis +5 vor. Daher entstehen zahlreiche Stickstoffverbindungen durch chemische, biochemische, geochemische und biogeochemische Prozesse.“ (Hanrahan, 2005)
Globale Stickstoffverbindungen und ausgewählte chemische Daten (Hanrahan, 2005)
| Oxidationsstufe | Verbindung | Funktionelle Anmerkungen |
|---|---|---|
| +5 | NO₃⁻ (Nitrat) | Starkes Oxidationsmittel; Ausgangspunkt der Pökelung (Reduktion zu NO₂⁻ / NO). |
| +5 | HNO₃ (g/aq) | Salpetersäure; industrielle Quelle für Nitrate. |
| +4 | NO₂ (g) | Braunes Gas; dimerisiert zu N₂O₄; Zwischenprodukt im Nitrit-/Nitratkreislauf. |
| +3 | HNO₂ (g/aq) | Salpetrige Säure; instabiler Vorläufer von Nitritsalzen (NO₂⁻). |
| +2 | NO (g) | Stickstoffmonoxid; zentrales Pökelmolekül. Bindet Myoglobin und stabilisiert die Farbe. |
| +1 | N₂O (g) | Lachgas; spielt nur eine Nebenrolle bei der Pökelung, ist aber Teil des Stickstoffkreislaufs. |
| 0 | N₂ (g) | Atmosphärischer Stickstoff; inertes Reservoir für Stickstofffixierung. |
| −3 | NH₃ (g) | Ammoniak; reduzierter Stickstoff, Ausgangspunkt der Nitrifikation. |
| −3 | NH₄⁺ (aq) | Ammoniumion; Substrat in mikrobiellen NO-Bildungswegen. |
| −3 | NH₄Cl (s) | Ammoniumchlorid; Vorstufe in historischen Pökelrezepten. |
| −3 | CH₃NH₂ (g) | Methylamin; weniger relevant für die Pökelung, zeigt reduzierte Stickstoffformen. |
Der Weg des Stickstoffs durch die Oxidationsstufen
Um die Chemie des Pökelns und die Rolle von Stickstoffmonoxid (NO) zu verstehen, lohnt es sich, den Elektronenfluss zu verfolgen, der Stickstoff durch verschiedene Oxidationsstufen führt. Diese Elektronenbewegung – Redox genannt (Reduktion–Oxidation) – erklärt, wie aus Nitrat (NO₃⁻) Nitrit (NO₂⁻) und schließlich NO wird, und warum NO sowohl in der Fleischverarbeitung als auch im menschlichen Körper so wirkungsvoll ist.
Man kann Oxidationsstufen als Maß dafür verstehen, wie „elektronenreich“ oder „elektronenarm“ ein Stickstoffatom ist. Bei −3 (z. B. NH₃) ist es voll mit Elektronen beladen. Bei +5 (z. B. NO₃⁻) ist es stark oxidiert, da Elektronen durch gebundene Sauerstoffatome abgezogen werden. Jede Stufe dazwischen repräsentiert eine Veränderung in der Elektronenbilanz.
Der Aufstieg von −3 zu +5
Ammoniak (NH₃, −3)
Vollständig reduziert; kein Sauerstoff gebunden, sehr elektronenreich.
Ausgangspunkt der mikrobiellen Nitrifikation, bei der NH₃ über Nitrit zu Nitrat oxidiert wird (Prosser, 1989).
Hydroxylamin (NH₂OH, −1)
Ein Sauerstoffatom wird gebunden, Elektronen werden teilweise vom Stickstoff abgezogen.
Zwischenstufe bei mikrobiellen oder industriellen Prozessen auf dem Weg zu Nitrit.
Lachgas (N₂O, +1)
Bereits teilweise oxidiert.
Nebenprodukt mikrobieller Prozesse im Boden; für das Pökeln kaum relevant.
Stickstoffmonoxid (NO, +2)
Das zentrale Molekül der Pökelung.
Bindet an das Eisen im Häm des Myoglobins, stabilisiert die rosa Farbe und verhindert Oxidation (Pegg & Shahidi, 2000; Honikel, 2008).
Entsteht durch Reduktion von Nitrit (NO₂⁻).
Nitrit (NO₂⁻, +3)
Das Arbeitstier der Pökelung.
Kann zu NO reduziert oder zu Nitrat oxidiert werden – je nach Elektronenfluss.
Nitrat (NO₃⁻, +5)
Die am stärksten oxidierte Form.
Stabil, in Pflanzen und Wasser weit verbreitet, Ausgangspunkt vieler Pökelsysteme.
Muss zunächst zu Nitrit reduziert werden, meist durch Bakterien.
Der Abstieg von +5 zurück zu −3
In der Pökelung konzentrieren wir uns auf den oberen Teil der Skala:
NO₃⁻ (+5) → NO₂⁻ (+3) → NO (+2)
Im lebenden Organismus wird dieser Weg weiter fortgesetzt – über N₂O bis hin zu NH₄⁺ oder NH₃, wodurch sich der Kreislauf schließt.
Dieser Zyklus ist nicht bloß ein chemisches Kuriosum:
- Er treibt die Zellatmung und den Energiehaushalt bei Bakterien.
- Er beeinflusst die Sauerstoffversorgung und mitochondriale Aktivität im menschlichen Gewebe (Lundberg et al., 2008).
- In der Pökelung steuert er Farbkonstanz, Aromaentwicklung und Haltbarkeit.
Redox in der Pökelchemie
Die Schritte Nitrat → Nitrit → NO lassen sich wie folgt zusammenfassen:
NO₃⁻ (+5) + 2 e⁻ → NO₂⁻ (+3) + O
NO₂⁻ (+3) + e⁻ → NO (+2) + O
In jedem Schritt werden dem Stickstoffatom Elektronen (e⁻) zugeführt – das ist Reduktion. Wenn wir sagen, „Nitrit wird zu NO reduziert“, meinen wir wörtlich, dass Elektronen in Richtung des Stickstoffatoms fließen, wodurch ein Sauerstoffatom abgespalten wird und NO entsteht.
Umgekehrt bedeutet Oxidation, dass NO Elektronen verliert und Sauerstoff aufnimmt – also wieder zu NO₂⁻ oder NO₃⁻ zurückkehrt.
Warum das wichtig ist
Zu verstehen, wo Stickstoff auf der Oxidationsstufenleiter von −3 bis +5 steht, ermöglicht es, sofort zu erkennen, ob eine Pökelreaktion (oder eine biologische Reaktion) oxidativ ist (aufsteigend, Elektronenverlust) oder reduktiv (absteigend, Elektronengewinn). Das ist entscheidend, weil:
- NO nur gebildet wird, wenn Stickstoffverbindungen reduziert werden, also Elektronen aufnehmen.
- Antioxidationssysteme in Pökellaken (z. B. Ascorbat) beschleunigen diese Reduktion, indem sie Elektronen spenden.
- Die Fähigkeit von NO, die Fleischfarbe zu stabilisieren, hängt damit zusammen, dass es Elektronen mit dem Häm-Eisen (Fe²⁺) teilt – und so die Oxidation zu Fe³⁺ (braunes Metmyoglobin) verhindert.
Kurze Zusammenfassung (in der vereinfachten Sauerstoff-Zählweise)
Wir haben bereits sehr viel gelernt – Zeit für eine kurze Wiederholung:
NOOO (NO₃⁻) → NOO (NO₂⁻) → NO
→ bei jedem Schritt wird ein Sauerstoffatom entfernt – das nennen wir hier Reduktion.
→ der umgekehrte Weg, bei dem Sauerstoff hinzugefügt wird, wird hier allgemein als Oxidation bezeichnet.
Nitrit (NO₂⁻) ist das reaktive Arbeitstier der Pökelchemie:
Es liegt chemisch nahe an NO, kann schnell reduziert werden und ist vor allem schnell, zuverlässig und steuerbar
(Pegg & Shahidi, 2000; Honikel, 2008; Sebranek & Bacus, 2007).
Oxidation und Reduktion anschaulich gemacht
Erweitern wir unsere Zusammenfassung mit einigen anschaulichen Darstellungen.
Der Nitrat–Nitrit–NO-Weg
Wir veranschaulichen dies mit einem einfachen Diagramm, das sowohl die Oxidation als auch die Reduktion von Nitrat – etwa aus Roter Bete – zeigt.

In der obigen Darstellung enthält Rote Bete Nitrat (NOOO / NO₃⁻)
Schritt 1: Nitrat verliert ein Sauerstoffatom (Reduktion) und wird zu Nitrit (NOO / NO₂⁻).
Dieser Schritt wird typischerweise durch Bakterien vermittelt, die das zusätzliche Sauerstoffatom für ihren Stoffwechsel nutzen.
Schritt 2: Nitrit verliert ein weiteres Sauerstoffatom und wird zu Stickstoffmonoxid (NO) – dem zentralen Molekül beim Pökeln.
Diese Schritte sind klassische Reduktionsreaktionen, da sie mit dem Verlust von Sauerstoffatomen einhergehen. Beim Pökeln bindet NO an das Häm-Eisen in Muskelproteinen (z. B. Myoglobin), stabilisiert die gewünschte rosa Farbe und verhindert die Oxidation zu braunem Metmyoglobin (Fe³⁺) (Pegg & Shahidi, 2000; Honikel, 2008).
Diese Reaktionen sind jedoch reversibel:
- NO kann ein Sauerstoffatom aufnehmen (Oxidation) und zu NO₂⁻ (Nitrit) werden.
- Nitrit kann ein weiteres Sauerstoffatom aufnehmen und zu NO₃⁻ (Nitrat) werden.
- In manchen Fällen nimmt NO direkt zwei Sauerstoffatome auf und wird zu NO₃⁻, ohne den Zwischenweg über Nitrit.
Dieses Wechselspiel zwischen Reduktion und Oxidation bildet die Grundlage des Stickstoffkreislaufs – sowohl beim Pökeln als auch im menschlichen Körper.
Biologische und chemische Wege zur Bildung von NO

Diese zweite Darstellung zeigt die beiden Hauptpfade der NO-Bildung:
Im Körper (enzymatischer Weg):
L-Arginin + O₂ → L-Citrullin + NO
Dieser Prozess wird durch die Enzymfamilie der Stickstoffmonoxidsynthasen (NOS) vermittelt, die NO für die zelluläre Signalübertragung, die Regulation des Blutflusses und das Immunsystem produzieren
(Lundberg, Weitzberg & Gladwin, 2008).
Außerhalb des Körpers (chemisch/bakterieller Weg)
Nitrat (NO₃⁻) wird zunächst durch Bakterien oder chemische Reaktionen zu Nitrit (NO₂⁻) reduziert.
Nitrit wird anschließend zu NO reduziert – typischerweise durch Reduktionsmittel (z. B. Ascorbat) in Pökellaken oder durch Hitze.
Beide Wege führen zu NO – chemisch identisch, ganz gleich ob durch menschliche Zellen, Bakterien oder Pökelsalze gebildet.
Erweiterte Erklärung von Oxidation und Reduktion im NO-Zyklus
(Bezieht sich auf die obige Abbildung – „NO-Zyklus“)
Diese dritte Darstellung verdeutlicht, wie Oxidation und Reduktion die Umwandlung zwischen NO₃⁻, NO₂⁻ und NO ermöglichen.
Reduktive Schritte:
NO₃⁻ → NO₂⁻ → NO (Verlust von Sauerstoffatomen, Gewinn von Elektronen)
Oxidative Schritte:
NO → NO₂⁻ → NO₃⁻ (Aufnahme von Sauerstoffatomen, Verlust von Elektronen)
Aus chemischer Sicht:
- NO₃⁻ (Oxidationsstufe +5) ist stark oxidiert und sehr stabil.
- NO₂⁻ (Oxidationsstufe +3) ist weniger stabil und kann leicht reduziert (zu NO) oder oxidiert (zu NO₃⁻) werden.
- NO (Oxidationsstufe +2) ist sehr reaktiv, kurzlebig und bindet direkt an das Häm-Eisen in Muskelproteinen beim Pökeln.
Ein Wandel im Verständnis
Die Fleischindustrie ist längst nicht mehr die einzige Branche, die die Bedeutung von Stickstoffmonoxid erkannt hat. Das Molekül, das über Jahrhunderte als gefährlich und „künstlich“ galt, besitzt Eigenschaften, die es heute in den Mittelpunkt moderner Forschung rücken. Ohne NO ist Leben nicht möglich – und doch wissen nur wenige davon, da seine Entdeckung erst in den 1980er- und 1990er-Jahren erfolgte.
Stickstoffmonoxid ist ein außergewöhnlich wichtiges Molekül.
Biologisch unverzichtbar
Vor einigen Jahren – bevor die Bedeutung von Stickstoffmonoxid erkannt wurde – betrachteten Verbraucher den Einsatz von Nitrit in Lebensmitteln (das sehr reaktiv und deutlich giftiger als Nitrat ist) mit großer Skepsis. Man verstand nicht, dass Stickstoff (N) in der Natur ganz leicht und häufig in NO (Stickstoffmonoxid), NOO (Nitrit) oder NOOO (Nitrat bzw. Salpeter) übergeht. Ebenso wird NOOO (Nitrat/Salpeter) oft und leicht zu NOO (Nitrit) und NO (Stickstoffmonoxid) umgewandelt. Wo man NO findet, ist es sehr wahrscheinlich, dass auch NOO und NOOO vorhanden sind – und umgekehrt. Das ist ein ganz normaler Bestandteil der Funktionsweise des menschlichen Körpers.
Dass Nitrit giftig ist, muss mit dem Hinweis ergänzt werden, dass Nitrit nur unter bestimmten Bedingungen toxisch wirkt. Welche Bedingungen das genau sind, wird ein zentraler Schwerpunkt unserer Untersuchung sein. Allein aus der Tatsache, dass eine Substanz unter bestimmten Umständen giftig ist, den Schluss zu ziehen, dass sie generell gefährlich ist und daher in Lebensmitteln nichts zu suchen habe, ist eine falsche Behauptung.
Im Laufe dieser Arbeit werde ich einen sehr wichtigen Vergleich einführen – nämlich zwischen Sauerstoff und Stickstoff. Sauerstoff ist dem Stickstoff darin ähnlich, dass er unter bestimmten Bedingungen toxisch ist und zum Tod führen kann. Tatsächlich kann man sagen, dass jede Zelle mit Zellkern im Rahmen ihres ganz normalen Stoffwechsels sowohl reaktive Sauerstoffspezies als auch reaktive Stickstoffspezies produziert (Griendling, 2016).
Wir wissen, dass selbst Sauerstoff unbeabsichtigte negative Folgen hat – etwa den Alterungsprozess beschleunigt und letztlich zum Tod des Körpers führt – obwohl er gleichzeitig die Grundlage allen Lebens auf der Erde bildet. Diese zweiseitige Wirkung begegnet uns auch im Bereich des Pökelns. Es ist von äußerster Bedeutung, diesen Mechanismus zu verstehen und verantwortungsvoll sicherzustellen, dass keine Bedingungen entstehen, unter denen Stickstoffverbindungen in irgendeiner Form dem Menschen schaden könnten.
Die bisher bekannten Fakten sind eindeutig: Stickstoffmonoxid (NO) – das zentrale Pökelmolekül – sowie seine „Verwandten“, Nitrat bzw. Salpeter (NOOO) und Nitrit (NOO), sind für das Leben von Menschen und Tieren und das Funktionieren unserer Körper unerlässlich. Stickstoff ist wahrscheinlich weder gefährlicher noch ungefährlicher als Sauerstoff.
Es ist in unserem Körper vorhanden!
Green et al. (1982) maßen die Konzentrationen von Nitrat (NO₃⁻) und Nitrit (NO₂⁻) in verschiedenen biologischen Flüssigkeiten und zeigten, dass diese Verbindungen ständig im menschlichen Körper vorhanden sind. Weit davon entfernt, „fremde“ oder toxische Zusatzstoffe zu sein, gehören sie ganz natürlich zur menschlichen Physiologie. Sie zirkulieren fortwährend durch Speichel, Blutplasma, Magensäfte, Urin – und sogar durch Muttermilch.
Das unterstreicht: Stickstoffmonoxid (NO), Nitrat (NO₃⁻) und Nitrit (NO₂⁻) sind untrennbare Bestandteile der menschlichen Biologie.
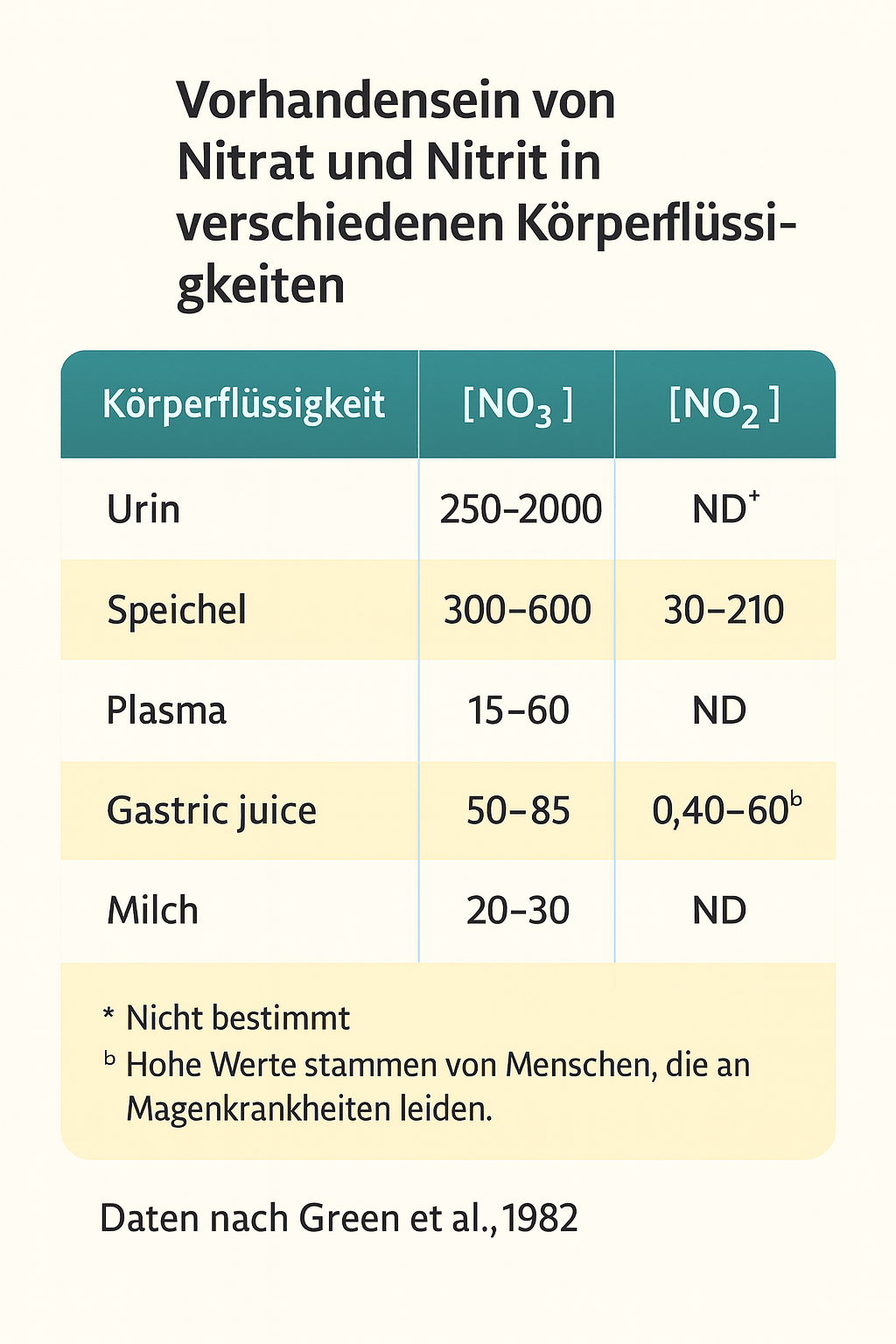
Wenn wir über Stickstoffmonoxid (NO) sprechen – ein essentielles Signalmolekül in unseren Blutgefäßen, unserem Immunsystem und in der zellulären Atmung – oder wenn wir Nitrat (NO₃⁻) und Nitrit (NO₂⁻) betrachten, ist es entscheidend zu erkennen, dass diese Moleküle nicht per se Toxine sind. Vielmehr handelt es sich um natürlich vorkommende Verbindungen, die für die Aufrechterhaltung normaler physiologischer Prozesse von zentraler Bedeutung sind. Sie übernehmen Funktionen wie:
- Kontrolle der Gefäßerweiterung (Vasodilatation),
- Regulierung des zellulären Sauerstoffgleichgewichts,
- Beteiligung an Immunreaktionen,
- Unterstützung der Bildung nützlicher Verbindungen in Speichel und Magensaft.
Deshalb sind pauschale Aussagen wie „Nitrit ist schädlich“ wissenschaftlich unbegründet, wenn sie außerhalb des Kontexts von Dosis und Balance getroffen werden (Lundberg & Weitzberg, 2010; Hord, Tang & Bryan, 2009).
Schlussfolgerung: Das Molekül, das Leben, Konservierung und Wahrnehmung verbindet
Stickstoffmonoxid ist nicht nur ein Pökelmittel. Es ist ein Molekül, das die Bereiche der Lebensmittelwissenschaft, der menschlichen Physiologie und der Umweltbiochemie miteinander verbindet. Seine Rolle im Fleisch ist der Einstieg in eine weitaus größere Geschichte – eine Geschichte, die die Regulation des Blutflusses in unseren Arterien, die Stabilisierung von Farbe und Geschmack in unseren Lebensmitteln, die Abwehrmechanismen unserer Immunzellen und sogar die Ernährung von Pflanzen im Boden umfasst.
In diesem Kapitel haben wir die Umwandlungen des Stickstoffs über seine verschiedenen Oxidationsstufen hinweg verfolgt. Von den reduzierten Ammoniumionen in verrottendem Material bis zu den hoch oxidierten Nitraten in Blattgemüse haben wir gesehen, wie Stickstoff durch das Leben in Zyklen von Elektronentransfer fließt. Im Fleisch wird dieser Fluss auf ein Ziel ausgerichtet: die Bildung von Stickstoffmonoxid – dem Molekül, das Farbe stabilisiert, Verderb verzögert und sensorische Eigenschaften bewahrt. Doch dieses gleiche Molekül, aus denselben Vorläufern gebildet, wirkt ebenso elegant und zielgerichtet in unserem Blut, unseren Lungen, unserem Gehirn und unserem Verdauungssystem.
Was einst als künstliches Konservierungsmittel gefürchtet wurde, ist in Wahrheit Teil eines fein abgestimmten und uralten biologischen Systems. Die Verteufelung von Nitrit und Nitrat in Lebensmitteln entspringt häufig einem unvollständigen Verständnis von Chemie und Biologie. In Wirklichkeit handelt es sich nicht um fremde Substanzen, die Lebensmitteln zugesetzt werden – es sind Moleküle, die in unserem Speichel entstehen, durch unser Blut zirkulieren und uns vor Infektionen und oxidativem Schaden schützen.
Pökeln ist daher kein Trick, der dem Fleisch künstlich auferlegt wird, um die Natur zu täuschen. Es ist eine Fortsetzung der natürlichen Chemie, die der Mensch sich zunutze macht, um Nahrung zu bewahren, Geschmack zu veredeln und den biologischen Rhythmen des Lebens gerecht zu werden. Wir pökeln Fleisch nicht einfach – wir nehmen teil an einem ununterbrochenen Zyklus molekularer Umwandlungen, der das Leben auf der Erde seit jeher erhält. Das ist nicht nur eine Lektion in Wissenschaft – es ist eine Lektion in Achtung. Pökeln ist keine künstliche Maßnahme; es ist ein Spiegelbild des Lebens selbst.

Literaturverzeichnis
Atkins, P., & de Paula, J. (2014). Atkins’ Physical Chemistry (10th ed.). Oxford University Press.
Beckman, J. S., & Koppenol, W. H. (1996). Nitric oxide, superoxide, and peroxynitrite: the good, the bad, and ugly. American Journal of Physiology.
Binkerd, E. F., & Kolari, O. E. (1975). The history and use of nitrate and nitrite in the curing of meat. Food and Cosmetics Toxicology, 13(6), 655–661.
Bogdan, C. (2015). Nitric oxide synthase in innate and adaptive immunity: an update. Trends in Immunology.
Brown, G. C., Cooper, C. E., & Wharton, D. C. (2001). Nitric oxide and mitochondrial respiration. Biochimica et Biophysica Acta.
Cassens, R. G. (1995). Nitrite-Cured Meat: A Food Safety Issue in Perspective. Food & Nutrition Press, Trumbull, CT.
Fang, F. C. (2004). Antimicrobial reactive oxygen and nitrogen species: concepts and controversies. Nature Reviews Microbiology.
Gladwin, M. T., et al. (2005). The emerging biology of the nitrite anion. Nature Chemical Biology.
Gladwin, M. T., Shelhamer, J. H., Schechter, A. N., Pease-Fye, M. E., Waclawiw, M. A., Panza, J. A., Ognibene, F. P., & Cannon, R. O. (2005). Role of circulating nitrite and S-nitrosohemoglobin in the regulation of regional blood flow in humans. Proceedings of the National Academy of Sciences, 97(21), 11482–11487.
Gray, J. I., Gomaa, E. A., & Buckley, D. J. (1996). Oxidative quality and shelf life of meats. Meat Science.
Green, L. C., Tannenbaum, S. R., & Goldman, P. (1982). Nitrate and nitrite in human saliva, gastric juice, plasma, and urine: dietary sources and endogenous synthesis. Science, 216(4542), 1131–1134.
Halliwell, B., & Gutteridge, J. M. C. (2015). Free Radicals in Biology and Medicine (5th ed.).
Hanrahan, G. (2005). Nitrate, Nitrite, and Nitric Oxide in Drinking Water. Royal Society of Chemistry, London.
Honikel, K. O. (2008). The use and control of nitrate and nitrite for the processing of meat products. Meat Science, 78(1–2), 68–76.
Hord, N. G., Tang, Y., & Bryan, N. S. (2009). Food sources of nitrates and nitrites: the physiologic context for potential health benefits. The American Journal of Clinical Nutrition, 90(1), 1–10.
Housecroft, C. E., & Sharpe, A. G. (2012). Inorganic Chemistry (4th ed.). Pearson.
Keilin, D. (1966). The History of Cell Respiration and Cytochrome.
Lehninger, A. L. (1975). Biochemistry: The Molecular Basis of Cell Structure and Function (2nd ed.). Worth Publishers.
Lundberg, J. O., Weitzberg, E., & Gladwin, M. T. (2008). The nitrate–nitrite–nitric oxide pathway in physiology and therapeutics. Nature Reviews Drug Discovery, 7, 156–167.
Lundberg, J. O., & Weitzberg, E. (2010). NO-synthase independent NO generation in mammals. Biochemical and Biophysical Research Communications, 396(1), 39–45.
Milton-Laskibar, I. (2021). Adapted from Niayakiru et al., 2020. Nitrate–nitrite–NO pathway [graphic source cited in-text; full reference needed].
Mitchell, P. (1961). Coupling of phosphorylation to electron and hydrogen transfer by a chemiosmotic type of mechanism. Nature.
Morita, H., Sakata, R., & Nagata, Y. (1997). Nitric oxide formation by Lactobacillus plantarum isolated from fermented foods. Journal of Bioscience and Bioengineering, 84(2), 165–169.
Nicholls, D. G., & Ferguson-Miller, S. (2013). Bioenergetics 4.
Otto, A., et al. (2010). Lightning-produced NOx and its role in the nitrogen cycle. Atmospheric Chemistry and Physics.
Pearson, A. M., & Gillett, T. A. (1996). Processed Meats (3rd ed.).
Pegg, R. B., & Shahidi, F. (2000). Nitrite Curing of Meat: The N-Nitrosamine Problem and Nitrite Alternatives. Food & Nutrition Press, Trumbull, CT.
Plant Nutrients and Lilies. (2010). University of Saskatchewan Extension Publications.
Prosser, J. I. (1989). Autotrophic nitrification in bacteria. Advances in Microbial Physiology, 30, 125–181. [or full citation if another source used]
Sebranek, J. G., & Bacus, J. N. (2007). Cured meat products without direct addition of nitrate or nitrite: what are the issues? Meat Science, 77(1), 136–147.
Shiva, S. (2013). Nitric oxide as a regulatory molecule in mitochondria. Biochimica et Biophysica Acta.
Tompkin, R. B. (2005). Nitrite. In P. M. Davidson, F. J. Montville, & A. L. Branen (Eds.), Antimicrobials in Food (3rd ed.). CRC Press.
Weitzberg, E., & Lundberg, J. O. (2010). Novel aspects of dietary nitrate and human health. Annual Review of Nutrition, 30, 189–213.